Genetics
From DNA to proteins
Also see: Biochemistry
The function of DNA within organisms is to codify the structure of proteins, according to the sequence of nucleotides (nitrogenous compounds anchored to a string of sugars and phosphates). Every group of nucleotides that codifies a particular protein is called gene, and the whole complex of genes found in an organism is called genome.
Protein synthesis occurs in two parts. In the transcription phase, the two strings that make up DNA are divided by an enzyme, and the sequence of nucleotides is copied by another enzyme in a single filament of messenger RNA (mRNA). Since each nucleotide is only coupled to another one, the sequence is copied accurately. There's an ecception: in retroviruses the genome is directly stored on the mRNA, and then transcribed on the host cell's DNA by reverse transcriptase.
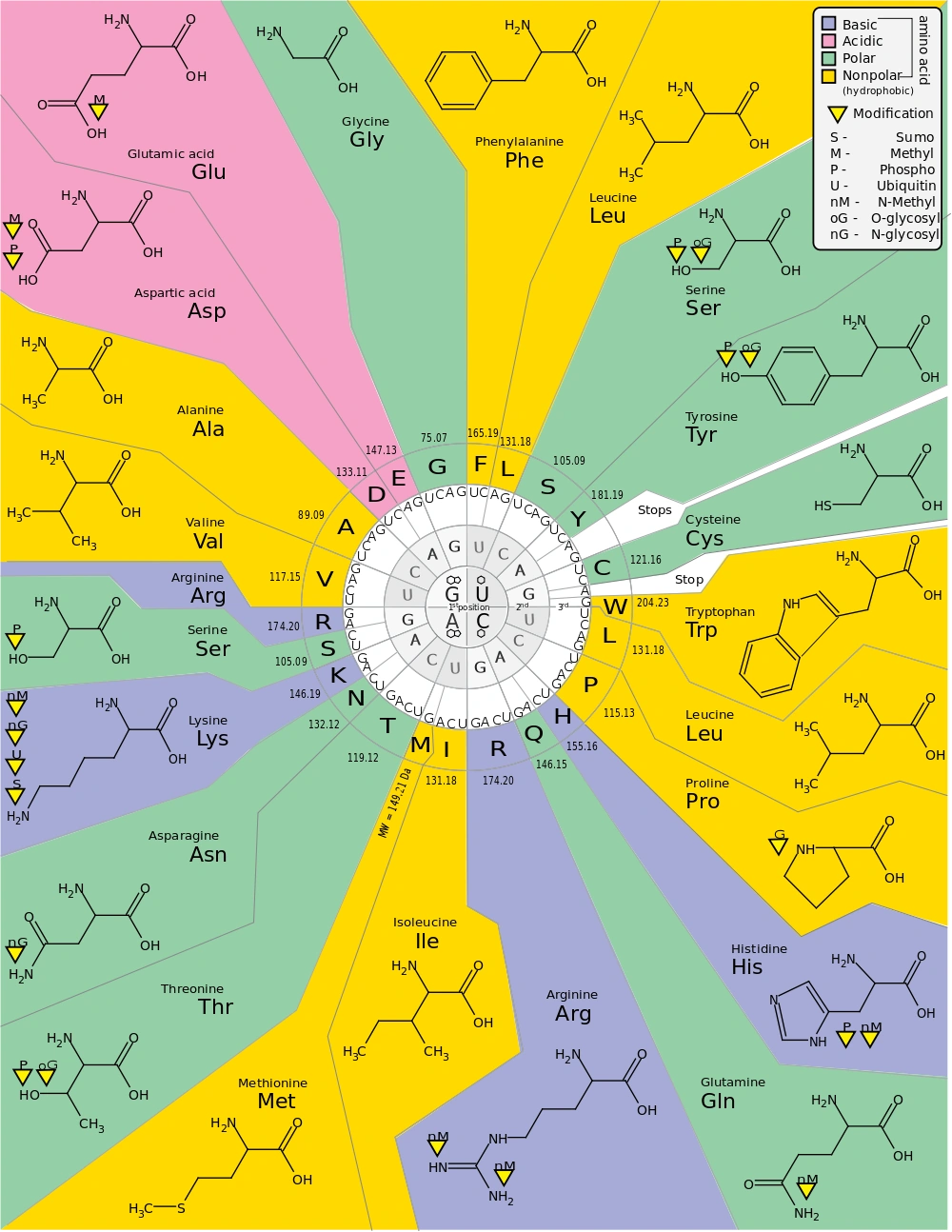
The genetic code, showing what amino acid corresponds to a particular codon. Most of this code is the same in every lifeform on Earth.
The sequence carried by mRNA reaches specialized organs, ribosomes, where starts the translation phase. Nucleotides are grouped in triplets, called codons: each codon codifies for a specific amino acid. Molecules of transport RNA (tRNA) anchor themselves to the codons of mRNA, each of them bound to an amino acid, and as the ribosome moves along the filament, they deposit a sequence of connected amino acids (polypeptides), until a particular sequence (UAA, UAG or UGA) signals the end of the process.
The polypeptide represents the primary structure of the protein, which then folds itself in helixes or laminae (secundary structure). These, in turn, fold themselves in the three-dimension tertiary structure. Two or more tertiary structures can, in some cases, be bound by hydrogen binds in a quaternary structure.
Genetic information
Genetic mutations
Genetic mutations are inheritable changes to the genome of an organism: by creating variation between the individuals of a population (together wiht processes such as meiosis and sex), they provide raw material for evolution. A mutation can be due to errors in replication and meiotic ricombination of DNA, or in the transcription and translation of mRNA, which in turn can be induced by chemicals or high-frequency radiations. Mutation are actually inherited only if they occur in the development of gametes or in the earliest stage of embryonal development.
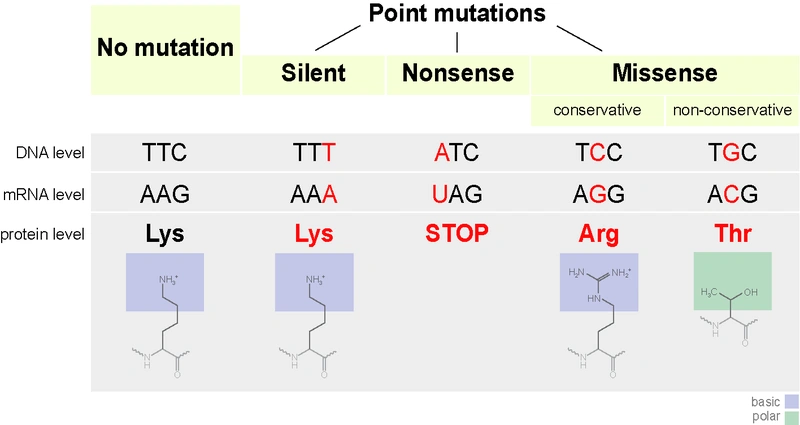
Examples of point mutations. The original triplet (TTC on the DNA, AAG on the mRNA, coding the aminoacid lysine) changes in TTT (also lysine: silent mutation), ATC (stop codon: nonsense mutation), TCC (arginine, same chemical properties: conservative missense mutation) and TGC (threonine, different chemical properties: non-conservative missense mutation).
According to their effect and scale, mutation are classified as:
- Small scale mutation, limited to brief sequences of nucleotides or genes.
- Point mutations, involving a single nucleotide. The most common type is an exchange between purins (A ↔ G) or between pyrimidins (T ↔ C).
- In silent mutations, the new codon codifies the same amino acid as before;
- In missense mutations, the new codon codifies a different amino acid;
- In nonsense mutations, the new codon is a stop triplet, codifying the end of the polypeptide.
- Deletions and insertions, in which one or more nucleotides are deleted or inserted in a sequence. If their number is not divisible by three, all the following codons are shifted and changed (frameshift mutation).
- Point mutations, involving a single nucleotide. The most common type is an exchange between purins (A ↔ G) or between pyrimidins (T ↔ C).
- Large scale mutations, extended to the structure of chromosomes: duplications, deletions of genes or genetic sequences, translocations of genes among non-homologous chromosomes and disappearance of alleles.
Heredity
The individual transmission of inheritable character so that the genetic variation produced by mutations and recombination can be preserved through the generations, rather than being watered down by hybridization. Since each organism in a species with two sexes receives half of its genome from the father and the other half from the mother, each gene is inherited by only one parent, and expressed as a whole. The two variations of the same gene, coming from the two parents, that codify for the same trait are called alleles (see here for more details and variations). This may not apply to asexual species.
An organism that has two identical alleles for a gene is called homozygote, and heterozygote if the gene exists in two different alleles. The transmission of traits to the offspring occurs according to the two Mendel's laws:
- Law of segregation: in two organisms, homozygotes relatively to a particular gene but in different way, different traits appear; for example, in the peas studied by Mendel, a purple flower (pp) and a white flower (ww). The offspring of these two organisms is a heterozygote (pw alleles), but rather than a mix of the parent's traits, only one is manifested: in this case, the colour purple. Thus, only one allele (the dominant one) is manifested, while the other one (recessive) remains inactive. Different sets of alleles (genotype) can codify for the same set of external traits (phenotype).
Crossing two heterozygotes (pw), the offspring can contain two phenotypes (purple and white) and three genotypes (pp, pw, ww) for each trait; on average it's composed for the 25% of homozygotes of a type (pp), for the 25% of homozygotes of the other type (ww) and for the 50% of heterozygotes (pw).
In some cases, the alleles can be incompletely dominant, and mix their traits: while homozygote snapdragons can be either white or red, a heterozygote hybrid can have pink flowers. In the case of codominance, the heterozygote phenotype can manifest both traits found in the parents: if a human has the blood group A and another has B, their offspring will have the blood group AB.
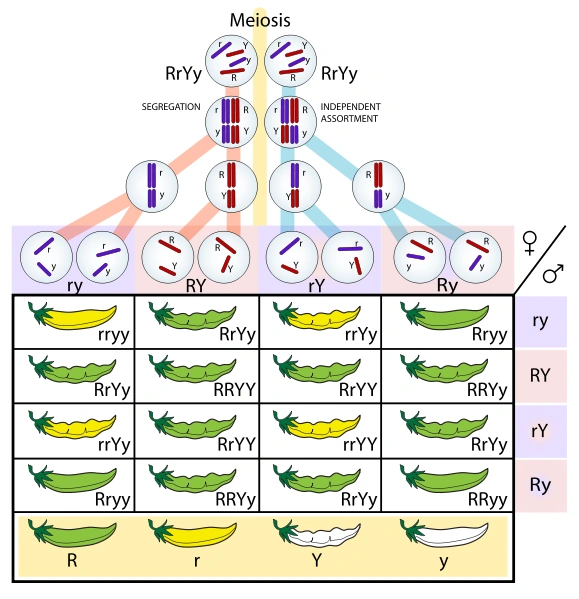
Mendel's law of independent assortment in peas. The "R" gene codifies the pod colour, the "Y" gene its shape: r (yellow) and y (smooth) are recessive alleles; R (green) and Y (rough) are dominant alleles. Each sex can produce four different gametes, with the various combinations of alleles (ry, RY, rY or Ry, in the red/purple bands of the table). As peas are diploid organisms, the table is a cross table with two variations.
- Law of independent assortment: when considering two traits at the same time, they're distributed among the offspring independently from each other, according to precise proportions. There is a 2:1:1 for both traits, which means that the four possible phenotypes (codified by sixteen genotypes) are found in a proportion 9:3:3:1, where, on average, for 16 children 9 show both dominant phenotypes, 3 the dominant phenotype for a trait and the recessive one for the other, 3 the opposite case and only 1 shows the recessive pheotype for both traits.
{kind=link}
{kind=link}
{kind=link}
The specific proportion given, of course, are valid only if the two alleles are equally common. According to the Hardy-Weinberg principle, given the two alleles of the same gene found within a population with the proportions p and q (where necessarily p+q=1), the distribution of the genotypes is (p+q)2=1, or p2+2pq+q2 =1. Here, p2 and q2 are the frequencies of the two homozygote genotypes, while 2pq is the frequency of the heterozygote genotype. For example, if the first allele is found in the 30% of the population and the other in the 70% (p=0.3, q=0.7), the frequencies will be 0.09 (pp homozygote), 0.49 (qq homozygote) and 0.42 (pq heterozygote).
Further generalizations are easy. When three alleles (p, q, r) are present, their frequencies will be (p+q+r)2=1, or p2+q2+r2+2pq+2pr+2qr=1;; if each individual is triploid and contains three alleles per gene, the frequencies will be (p+q)3=1, or p3+3p2q+3pq2+q3=1. As a general rule, the distribution of genotypes will be (a1+a2...+an)p, where n is the number of alleles and p the ploidy number. This principle assumes both a random mating and strict mendelian inheritance law, which do not always apply.
Epigenetics
Natural selection
Another factor fundamental for evolution is the competition for resources, necessary consequence of the exponential growth of a population that forces it, sooner or later, to exceed the carrying capacity of the environment (Malthus principle). Thus, only a small fraction of the conceived genotypes manages to be passed to the next generation. Natural selection is the process in which the genetic traits that support their own survival and multiplication (faster reproduction, higher energetic efficiency, improved sense organs or natural weapons, cooperation, etc.) tend to increase in number faster than the others, so that the average genotype in the population becomes more and more fit to its environment.
Natural selection is stabilizing when it keeps a trait around an average value, being the extreme values less beneficial. It's disruptive when it favours extreme values but not average ones: for example, in salmon males, which can mate easily if they're large and strong ("hooknose") or if they're small and furtive ("jack") but not if they have an average size, which has neither strength nor stealth. Finally natural selection is directional when it favours an extreme rather than the other one, so that this value becomes the new average: in this way, the whole population gradually changes its genetic traits. Both directional and disruptive selection can induce speciation: the first one by changing the whole population, the second one by splitting it in two species.
The group of traits favoured by natural selection is called fitness. The fitness of a particular genotype is measured as the ratio between the number of individuals that carry it in a given generation and that number in the previous generation: if the ratio is greater than 1, the genotype increases in frequency.
Genetic flux and drift
Not all genotypic variations are induced by the environment. The genetic flux is the relocation of genes or alleles towards the inside or the outside of a population due to migration (even if accidental) of a group of individuals: the species that disperse their seeds or gametes in the environment are particulary suscetible to genetic flux. In a small population, random events that change the reproductive success of a few individuals, carrying a certain gene or allele, can influence its overall distribution altering the Hardy-Weinberg equilibrium: this phenomeon is called genetic drift.
A small population, separated from a larger one, can have a distribution of genotypes very different from the original one, simply because of random variety, and it will matain it with its expansion: this is known as founder effect when the new population is produced by a miration, and bottleneck when it's caused by a catastrophe (this is the cause of the low genetic diversity of cheetahs and elephant seals, or, in the case of humans, a consequence of the eruption of Toba 74 000 years ago). Such events can completely remove even common alleles, or greatly change their distribution: the reproductive isolation of askhenazi jews has caused a high incidence of the Tay-Sachs disease.
The mechanism called Muller's ratchet is typical of asexual reproduction: since without ricombination the entire genome is transmitted as a whole, defects due to harmful mutations pile up through generation, as they can't be compensated by the "healthy" alleles of another parent. This increases the genetic load, that is, the reduction of fitness due to genotype imperfections (it's calculated as L=(w-v)w, where w is the maximum fitness in the population and v the average one).
Sexual selection
Since tha main units of natural selections are not individuals as much as genes, often the individual survival is subordinate to the transmission of genes. When a species with sexual reproduction (that is, when more than one individuals are needed for procreation), there's often a specialization of gametes (see here), the largest and less numerous gametes (eggs) become a scarce resource which forces the members of thew other sex(es) into competition to fertilize them. This sex, generally identified as female, is the limiting sex.
When this happens, the competing sex undergoes adaptation that can reduce their chance of individual survival, but improve the chance of reproduction. Male competition often occurs as lekking, that is, showing ornamentation, fighting with other individuals, or both (for example, the tail feathers of peacocks, or deer antlers). There's also spermatic competition: males can employ spermicide chemicals that kill the competitors' gametes, or physically block the female genitals (with their own torn genitals, in bees and many spiders), or simply producing gametes in larger quantity or better quality. It has been observed, in closely related mammal species, that the more promiscuous species (e.g. humans and chimpanzees) have larger testicles than specie where each female mate with only one male (e.g. gorillas).


{kind=link}
The diopsid fly Teleopsis dalmanni, with developed eyestalks as ornamentation. The reproductive advantage they confer overcomes the difficulties they cause.
Often, the results of sexual selection can actually hinder the life of the competing sex: the vivid coloration and the cumbersome tail of male peacocks and paradise birds make them easier preys, while the enlarged eyestalks of the Diopsidae flies harm their sight. According to the handicap hypothesis, this actually makes them even more successful in reproduction, whatever the cost to their personal life, since they prove to have good genes by surviving despite having such handicaps.
Kin selection
Since related individuals within a population share a fraction of their genes, natural selection can lead an organism to disregard its own survival/reproductive success in order to benefit its relatives, as long as the total advantage received by the gene(s) for altruistic behaviour (which is also found in the beneficiaries of altruism) is greater than the harm received by the altruistic individual.
According to Hamilton's rule, such a behaviour is selected for if rB>C, where C and B are the total cost (for the acting individual) and the total benefit (for the beneficiaries) of the action, both measured in the same unit, and r is the degree of relatedness between the individuals. For example, r=1 for clones and identical twins, ½ for parents, children or siblings, ¼ for grandparents, uncles and aunts, first cousins and grandchildren, etc. From an evolutionary point of view, the survival of a particular individual is (for it) worth as much as the survival of an identical twin, two children/siblings, four grandchildren/ cousins, etc.
A 2011 study has tested Hamilton's Rule through robots, discovering that a distinctly altruistic behaviour could arise in as few as 500 generations. Thus, in social species, relatedness can bring to the altruistic sharing of resources (for example, blood in vampire bat colonies). Kin selection is especially effective when organisms can easily recognize their relatives, or at least the other carriers of the self-helping gene (the "green-beard effect"), but in "viscous populations" (low dispersion and thus strong genetic mixture) even indiscriminate altruism can be selected for.
Speciation
Speciation is the formation of a new species due to the gradual accumulation of modifications that eventually prevent the two populations from hybridizing (reproductive isolation). The mechanisms that make impossible for two individuals from these populations to breed can be pre-zygotic (incompatible visual, auditory or chemical signals, different times and places of reproduction, lack of anatomical compatibility, inability of the gametes to meet) and post-zygotic (zygote unable to develop, weakness or sterility of hybrids). The mechanisms become more and more radical (for example, first the two populations can have less fit hybrids, then the hybrids become sterile, then they can't be born at all, then fecundation can not occur, etc.) until the two populations don't recognize each other anymore as reproductive partners, and become two distinct species.
(Note that the line between species is often blurry, since speciation is constantly in progress; also, the reproductive definition of species can't apply to asexual organisms, where the classification in species is more arbitrary.)
{kind=link}
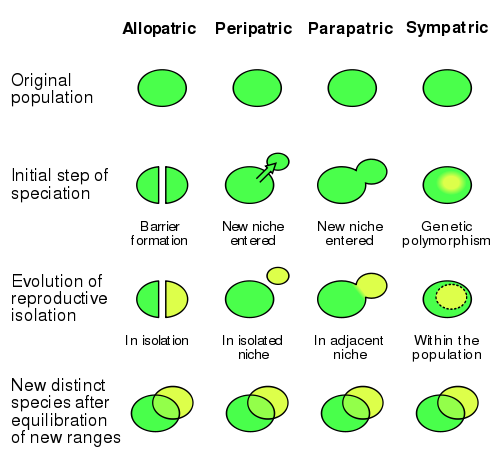
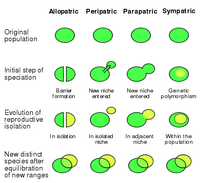
Scheme of the four modes of speciation, in three phases of the process.
- In allopatric speciation, parts of a population are separated by a physical barrier such as a river or a mountain range: in an ecological sense, isolated environments can be mountain peaks, oasis in a desert, host populations for a parasite, etc. With time, genetic drift and possibly natural selection if the new environments diverge enough, the two populations accumulate differences until they're not able to interbreed anymore.
- Peripatric speciation is similar to allopatric speciation, combined with a bottleneck: a small part of the population separates from the rest and colonizes a new environment.
- In parapatric speciation, a species develops ecotypes, that is, different phenotypes adapted to slightly different conditions in adjacent areas, until drift and selection turn the ecotypes in distinct species.
- In sympatric speciation, intraspecific competition causes the specialization of different coexisting populations to exploit different resources, or the same resources in different ways; this produces variation and inhibits interbreeding, once again causing speciation.
Often species with a wide geographical extension, living in different climates, develop chains of ecotypes called clines. The adjacent ecotypes can usually interbreed, but those at the extremes of the chain cannot due to the cumulative genetic differences: the extinction of the intermediate links can cause the complete and definitive separation of the extreme ones as distinct species. Clines are called ring species if the ends of the chain live close to each other, possibly occupying different niches (for example, the populations of gulls around the North Pole).
Laws of evolution
Evolution is not a deterministic system: while there are equations that describe some phenomena, such as the Hardy-Weinberg equilibrium, there's no way to predict the outcome of evolution from given start conditions. It's possible, though, to recognise long-term trends that make certain developments more likely than others, and other ones impossible.
Laws on the global pace of evolution:
- Dollo's Law (irreversibility of evolution): an organism, population or species cannot return to a previous condition already realized in its ancestors. While atavic traits might emerge as malformations if the genes that codify them are still present, a structure completely lost in evolution, for example the gills for land vertebrates, can't re-emerge in the same form that was lost, even if it could be beneficial. Of course, there's the possibility of a new development similar to the other (some turtles developed structures to absorb oxygen in their rectum, which could form gill-like organs).
- Roulliet's Law (complication of systems): the development of organisms, clades or other biological systems makes their organization more and more complex with time, increasing the number and the diversity of their sub-systems. The first lobopods (likely ancestors of arthropods) were probably similar to caterpillars or velvet worms, with a number of simple appendages which, in time, specialized into legs, maxillae, antennae, palps, claws, etc. In the same way, the complexity of biomes and ecosystems tends to increase, with a greater and greater number of species involved and connections between them.
Williston corollary: organisms tend to have less and less limbs and segments, but more specialized ones. - Schmalhausen's Law (greater integration): biological systems become more and more integrated. As complexity increases, the specialization and reduction in number of parts means that each of them can perform less functions, and thus each becomes more and more irreplaceable. For example, a sponge ground into mush can reconstruct its whole body structure, being so simple; a flatworm and a starfish can regrow large sections of their bodies; a lizard can regrow its tail, but not the more complex limbs or head. A cockroach, whose nervous system is distributed all around the body, can survive for a long time with its headcut off, but a vertebrate, whose nervous system is concentrated in the brain, cannot.
- Law of Limitless Progress: as long as there is life, there is evolution. Evolutionary development, or any other form of progressive change in biological systems, does not end if the basic properties of life (metabolism, competition for limited resources, imperfect remplication) are still present.
- Law of Acceleration: as complexity increases, the average longevity of a species or clade decreases (the individual lifespan might increase, though), and the rate of evolution per time unit increases. This is probably due to the fact that a more complex organism (a protist with a cellular nucleus rather than a bacterium, a multicellular organism rather than a single-celled protist, an animal or plant with tissues and organs rather than undifferentiated mats of cells) has more possibilities to evolve because it has more structures available to change. It's also possible that the ability to evolve, itself, evolves, producing conditions that make evolutinary change easier and faster (DNA, nucleus, sexual reproduction, gametes, HOX genes, etc.)
Schmalhausen corollary: organisms with a higher trophic level evolve faster. In his classification, the order of speed (from the slowest evolution to the quickest) is: preys surviving thanks to the high population → preys that survive thanks to passive defenses (shells, thorns, poison, etc.) → preys that survive by fleeing or fighting → predators. - Law of Genetic Variety: everything that is alive is genetically various, and tends to increase its variety with time. Hybridization does not occur in nature, except at the level of species: two orders/classes/phyla always split in two, and two never merge in one. There are cases of extreme symbiosis, such as the migration of bacteria in early eukaryotes as mitochondria and chloroplasts, or the association of algae and fungi in lichens; but such events still increase diversity, as they create a third new group. Every group of species, large as it may be, always traces back its ancestry to a single species.
- Law of Ecological Orientation: evolution does not have a final goal; the features of the organisms are always shaped by their environment, but also constrained by the internal development processes - so that the same environmental obstacle can be overcome with different adaptations. On Sivatherium, Pavel Volkov gives as example several species of fish that live in porrly oxygenated waters: everyone of them is adapted to absorb more oxygen through a specialized structure, but this structure is the mucouse membrane of the mouth in electric eels and of the intenstine in mud loaches, air bladders opening on the gills in a catfish, etc.
- Eldredge-Gould's Law (punctuated equilibria): evolutionary development, while always gradual, is not continuous: significant features often arise in a relatively brief timespan, and then remain unchanged for a much longer period. Volkov gives examples from the evolution of plants: photosynthesis, multicellularity, spores, organs and tissues, secundary growth, seeds and flowers. Each major change is followed by a large radiation of species (in the case of plants, the flourishing of unicellular algae, seaweed, mosses, ferns and clubmosses, early trees, conifers and flower plants). Of course, the "relativly brief timespan" is still in the order of hundreds of thousands or millions of years.
- Law of Constant Number of Species: as long as the conditions of the biosphere are stable, the rate of speciation and extinction is roughly equal, keeping the total number of species costant. Of course, counting mass extinctions, stellar evolution, the expansion into new niches such as dry land in the early Paleozoic, etc. it cannot be said that the whole biosphere is ever really stable, so this principle is really only relevant on a smaller spatial and temporal scale.
- Lotka's Law: the total biomass of an entire system increases with time as long as resources are available.
Laws on the development and extinction of clades:
- Principle of Exaptation (or pre-adaptation): often, a preexisting trait allows the organism to expand in a new niche by performing a different function than it had when it first appeared. For example, the ancestors of birds (theropod dinosaurs) developed feathers for thermic insulation, then (probably) for display, and finally they used them for flight. When the first tetrapods reached land, they already had lungs, fleshy fins, efficient kidneys etc. developed in water for other reasons than leaving water; other vertebrates, such as sharks and ray-finned fish, that lacked these pre-adaptations could not accomplish the same.
- Law of Equivalence of Conditions: all conditions of an environment are necessary for life, and if even one parameter falls outside the tolerance of a species, this will disappear.
- Cope's Rule: new groups of organisms evolve from the less specialized members of the groups that precede them. For example, the first representatives of mammals were mouse-like or shrew-like omnivores and insectivores able to survive in awide range of environment, and they in turn descend from the small cynodonts that survived the Permian Extinction, whereas their close parents gorgonopsians, being large and specialized, disappeared. In the same way, when considering future evolution, we can know that the most specialized forms of today are the most likely to disappear, and predict that they will be replaced by the newly specialized descendants of today's generalist organisms.
- Darwin's Rule (divergence)/Osborne's Rule (adaptive radiation): as a group develops through time, it divides in a greater and greater number of sub-groups with different specializations. Starting from the small, unspecialized forms as per Cope's Rule, in less than 20 million years mammals produced specialized predators or browsers, specialized flying, swimming and tree-dwelling forms; but of course each new specialization was realized in a different lineage, producing a wide and relatively quick radiation from the common ancestor. This is due to the adaptation to different niches, in order to reduce competition.
- Deperet's Rule (progressive specialization): when a group starts to specialize, it can only specialize further and never return to a more generalized state. This occurs because individuals born with less specialistic features will be outcompeted and won't have an advantage in any environment. Often, specializing groups also increase in size and weight, though some adaptations (e.g. burrowing, flying or parasitic lifestyle) might produce the opposite effect.
- Marsh's Rule: more specialized forms are more likely to become extinct, as they have a diminished ability to adapt to changing conditions. It's a direct consequence of Deperet's Rule (every change away from their particular specialization will make them less fit for their environment) and Dollo's Law (past specializations cannot be undone, and they leave traces on every future development).
Laws on the features of organisms:
- Principle of Ecological Conformity: the morphology (form), physiology (working) and behaviour of an organism are direct products of the environment it lives in, and they reflect its features.
- Rule of Conformity of the Environment to the Genetics: the existence of every organism is a careful balance between the external conditions and the internal constraints of its structure, so that it can exist only when the environmental conditions do not exceed the tolerable limits imposed by those constraints. For example, if all the trees in the african savanna were to disappear, every browser would have to adapt to grazing, but giraffes wouldn't be able to do so as they have a neck specialized for treetops browsing, which they couldn't eliminate (see Dollo's Law and Marsh's Rule).
- Bergmann's Rule and Allen's Rule: in a group of closely related species that live in different biomes, those that live in the colder ones will be larger than those that live in warmer ones (Bergmann), and they will have shorter limbs/appendages (Allen). This happens because large size and reduced appendages limit heat loss (see here); examples of these rule are canids (arctic foxes and wolves compared to their relatives of temperate areas), elephants (woolly mammoths compared to savanna elephants), bears (kodiak and polar bear compared to sun bears and sloth bears) and even humans (Neanderthals compared to H. sapiens). "Abyssal gigantism" observed in deep-sea crustaceans and molluisks is probably a consequence of Bergmann's Rule.
Hesse corollary: the colder-climate forms also have, in proportion, larger hearts. - Gloger's Rule: organisms that live in warm and wet habitats are usually more pigmented, while those that live in colder and/or drier habitats have plain colours. Heavy pigmentation protects the body from excess radiation, while drab colours help in cryptic camouflage in a bare environment; among birds, it has been observed that feather pigmentation protects them from bacteria thriving in wet conditions. There is an exception in humans: a relatively dark skin is found in tibetan people, to protect them from UV light reflected from the ice, and inuit people, whose diet rich in vitamin D allows them to survive with a lesser absorption of light.
- Rensch's Rule: if a species has sexual dimorphism (that is, the sexes look different), the difference in size increases with the absolute size if the male is larger, while decreases in the opposite case. This means that a species with a strong sexual dimorphism can have either a large female and a huge male (e.g. elephant seals) or a small female and a tiny male (e.g. anglerfish, green spoonworms), but not, say, a large male and a huge female or a small male and a tiny female. The reason for such a condition is unknown, but Rensch's Rule has been confirmed by many observations.
Laws on ecological niches:
(see here)
- Laws of Niche Restriction:
- Species must be specialized for a particular niche in order to survive.
- Ecological niches tend to narrow in order to increase the chance of mating: asexual species have wider niches than sexual species (as they need less this effect), and species with a sparse population or incapable of moving have narrower niches.
- Laws of Niche Segregation:
- Selection in relatively empty spaces leads to segregation.
- Niches are segregated becaus of competition between species (as competition decreases if the species specialize to consume different resources).
- The niches of closely related species are segregated to limit crossbreeding (as a hybrid of two species with slighlty different niches will be less fit than its local parent species in either niche).
Consequences of evolution
Nested hierarchies
See main article: Phylogeny
Biogeography
Homology and analogy
Vestigial structures
References
- Berkeley's Evolution 101, a simple and extensive guide to evolution
- A summary of the mechanism of DNA replication and protein synthesis
- Animal Genome Size Database