| Part of a series on speculative biology |
|---|
|
Champion speciators |
On the largest scale, the shape and the basic structure of organisms are dictated by two things: their function and mechanics. The size of a living being, the way organs and cavities are arranged inside the body, the way the parts of the skeleton interact are also the basis for any further speculation about its physiology.
Body shape[]
Cells and tissues[]
Symmetry[]
Symmetry is extremely important for the organization of the body: it provides a regular disposition of the bodyparts and often redundancy of limbs and organs. Most of pluricellular organisms on Earth show some form of symmetry, in four distinct degrees:
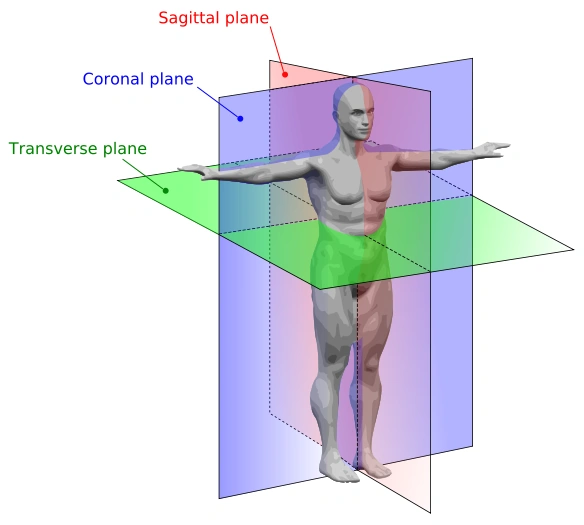
The three anatomical planes, only one of which divide in two specular halves a bilaterally symmetrical organism.
- Asymmetry is very rare, at least between moving organisms, since it's a very inefficient way to organize the body and, if specialized organs are present, doesn't offer any reliable backup in the case of damage. Some primitive pluricellular organisms are completely asymmetrical, such as sponges and placozoans.
- Spherical symmetry: the organism can be divided in two specular halves by each of the three anatomical planes (sagittal, coronal and transverse, see image on the right). It's found in organisms that live in a 3-dimensional environment, submerged in a fluid dense enough to counteract gravity, and therefore they don't have any reason to privilege a specific direction: microscopic protozoans such as radiolarians, heliozoans and Volvox.
- Radial symmetry: the organism can be divided in specular halves by a number of planes (sagittal, coronal and possibly other) that pass through the dorsoventral axis. It's common among animals that live under the strain of gravity but motionless, and therefore distinguish only a dorsal side from a ventral one (eg., corals and sea anemones). The ventral side of a creature will produce possible limbs and will protect the most precious organs, such as the genitals, while the dorsal side will concentrate the passive defenses (shells, thorns, bony plates)[1]. With the exception of jellyfish, that move in the direction of their back anyway, radial animals never move much; the only bilateral animals subsequently turned radial are the echinoderms (e.g. sea urchins and starfish). Common forms of radial symmetry include:
- trimerism (body divided in three symmetrical parts) in some radiolars and in the extinct trilobozoans;
- tetramerism (four parts) in some jellyfish;
- hexamerism (six parts) and octamerism (eight parts) in corals (respectively Hexacorallia and Octocorallia);
- biradial symmetry, used in the Ctenophora phylum: the body is divided in alternate quarters, each of which is different from the neighbouring ones but identical to the opposite one;
- pentamerism (five parts), the most common one: the symmetry axis are placed at 72° from each other. All the echinoderms show this symmetry, as well as many flowers and fruits, for example the petals of the dog rose and the seeds in an apple.
- Radial flowers (such as the hexameric lily or the pentameric buttercup) are called actinomorphic.
- Bilateral symmetry: the organism can be divided in specular halves only by the sagittal plane. Animals that move in a specific direction rather than stay fixed on the ground/seafloor will develop an anterior extremity (a "head") that will meet first the new object and will interact with them. It will therefore contain a mouth, the main sense organs (and the central nervous system associated to them, if it exists), and perhaps simple manipulation organs such as tentacles. The right and the left side should remain specular, with few exceptions, since they have no reason to diversify: in fact, an asymmetry of limbs or muscles could cause the creature to move in circle rather than forward. Almost all animals on Earth, be they earthworms, wasps or turtles, are bilateral. Bilateral flowers, such as orchids and Lamiales, are called zygomorphic.
- While most plants are asymmetrical on the whole, they often have branches or leaves arranged as a helix around the stem (helical growth); the cells of the Phycomyces fungi and the seaweed Nitella are helix-shaped, probably due to the arrangement of fibres in the cell wall.
| Location term 1 | Location term 2 | Axes | Meaning |
|---|---|---|---|
| Superficial | Deep | - | Closer to the surface/the centre |
| Anterior | Posterior | Anteroposterior | Closer to the front/the back |
| Medial | Lateral/peripheral | Mediolateral | Closer to/farther from a symmetry axis |
| Dorsal | Ventral | Dorsoventral | Closer to the spine/the stomach |
| Proximal | Distal | Proximodistal | Closer to the base/the tip of an appendage |
| Rostral/cranial/cephalic | Caudal | Craniocaudal | Closer to the head/the tail |
| Oral | Aboral | Oral-aboral | On the same/opposite side of the mouth |
| - | - | Longitudinal | Longest symmetry axis of the body |
| - | - | Transverse | Axis perpendicular to the longitudinal one |
Note: some terms of location may be synonimous or meaningless depending on the shape and symmetry of the body (for example, human upright posture makes dorsal/ventral synonimous to anterior/posterior).
Repetition and specialization[]
In a primitive cell colony, different parts will tend to specialize to different functions, allowing them to maximize their efficiency - but they won't be able to survive on their own anymore, turning the colony in an organism. Ancestral organisms tend to have more repetition in their structure:
Roulliet Law: The development of organisms, clades and other biological systems makes their organization more and more complex, with the differentiation and specialization of their sub-systems.
Williston corollary: Organisms tend to have less limbs and segments than their ancestors, but these limbs and segments will be more specialized.
Metamerism is the repetition of body segments with similar or identical structure, called somites or metameres in animals, and phytomers or metamers in plants (it's something different from mere segmentation, as in tapeworms, which is only a characteristic of external tissues). Annelids such as leeches and earthworms show homonomous metamery: all the segments are almost identical to the others, and they contain muscles, nephrids (excretion organs), part of the nervous chord and the digestive tract, a complete reproductive system and sometimes parapods (appendices without articulations). Tagmatization is the fusion of more segment in specialized functional group (tagmata).
Arthropods are instead an example of heteronomous metamery, that typically arises from the specialization of homonomous metamery: each segment is very different from the others, and so are the limbs, that become legs, claws, chelicerae, palps and jaws. The head of insects, crustaceans and myriapods is made up by five fused segments, the head of trilobites and Chelicerata by seven; today, the body of insects appears composed by only three tagmata (head, thorax and abdomen), the body of many arachnids only by two (cephalothorax and abdomen); scorpions and crabs have a large cephalothorax and many abdominal segments, while mites appear to be entirely devoid of segmentation.
Even when organs are specialized in function, they might have a very generic structure. Systems appear at first as a diffused net (of nerves, blood vessels, etc.) around the body, usually developing a greater centralization, with one or few main organs (brain(s), heart(s), etc.). The development "diffusion of chemicals from the environment" → "diffuse net that carries chemicals around the body" → "centralized system that controls the net" happens usually as a function of size (see the following paragraph).
Furthermore, while an efficient design might require every function to be executed by an organ, the convoluted evolutionary story of living beings often cause an organ to execute many functions. For example, the pharynx of terrestrial vertebrates is both part of the digestive and respiratory system; jellyfish and flatworms eat and excrete waste through the same hole; sea cucumbers and turtles breath through the rectum, while the chinese soft-shelled turtle also urinates through the mouth; in reptiles and birds, the digestive, urinary and reproductive tracts all open in the cloaca.
Conversely, organisms could execute the same function with different organs: squids have two hearts, octopuses have three, while ruminants have up to four stomachs. Finally, some organisms might forsake an organ completely: antlions don't have a stomach, because they eat only food digested by their enzymes outside their body, while many parasite worms don't have an intestine either, because they absorb only the nutrients already digested by the host.
Volume and surface[]
The relation between the volume and the surface of a solid is extremly important in several fields of biology. While the volume grows in proportion with the cube of the linear dimension, the surface grows in proportion only with the square, and therefore as a solid grows larger and larger its surface becomes (in relation to the volume) smaller and smaller:
| Side | Surface (cube) | Volume (cube) | Area:volume |
|---|---|---|---|
| 0.1 | 0.06 | 0.001 | 60 |
| 0.2 | 0.24 | 0.008 | 30 |
| 0.5 | 1.5 | 0.125 | 12 |
| 1 | 6 | 1 | 6 |
| 2 | 24 | 8 | 3 |
| 5 | 150 | 125 | 1.2 |
| 10 | 600 | 1000 | 0.6 |
| 20 | 2400 | 8000 | 0.3 |
| 50 | 15 000 | 125 000 | 0.12 |
That means that very small organisms have a very large surface in relation to the volume. Since mass, weight and metabolic activity (of every part of the body, and the whole) are proportional to the volume, while surface areas and cross sections are proportional to the surface, unicellular organisms can simply absorb the needed chemicals through the surface, and that's enough to sustain their processes; for larger organisms, the surface is too small, and they need complex systems to distribute the chemicals around the bodyparts.
- Functions proportional to area: absorption of chemicals and light, heat dispersion, strength of muscle and bone, brain activity, amount of fluids delivered to the tissues, air/water friction, drag and lift
- Functions proportional to volume: mass and weight, retention of heat, metabolic energy production, storage of chemicals, muscular power, blood pressure
When a large-sized organism need functions proportional to a surface, it can increase its surface but not its volume through wrinkles, laminae, gyri, branches: pulmonary alveoli, intestinal villi, capillaries, leaves, gills are all structures that allow to efficiently absorb or release chemicals (or light, in the case of leaves) without unnecessay bulk. In the same way, brain gyri extend the thin cerebral cortex, and the mitochondria ridges permit a faster oxidation of glucose.
Small organisms easily disperse heat - something very dangerous for small mammals and birds, that produce their own heat through metabolic activity, forcing them to stay much larger than cold-blooded animals - while large ones tend to retain it, and thus in a hot climate they need expanded surfaces, such as the ears of elephants.
Another, more evident consequence is that as an organism increases in size it needs proportionally thicker and stouter supports: thus, elephants and sauropods have pillar-like limbs while insects and spiders walk on such thin legs. If a mouse is increased in size a hundred times from 5 cm to 5 m, it becomes 1003 = 1 000 000 times heavier, while its bones are only 1002 = 10 000 stronger: it's therefore a hundred times heavier than its skeleton can bear and it will need a radically different skeletal structure.
| Material | Strength |
|---|---|
| Muscle | 10 atm |
| Wet chitin | 200 atm |
| Silica | 300 atm |
| Dry chitin | 350 atm |
| Cartilage | 350 atm |
| Soft wood | 380 atm |
| Hard wood | 500 atm |
| Tendon | 1000 atm |
| Bone | 1700 atm |
| Dentin | 3000 atm |
Note: A pressure of 1 atm equals roughly the weigh of 1 kg, in Earth gravity, on each square centimetre.
Many other peculiar features of small-scaled life are explained by the large area:volume ratio:
- insects can carry loads hundreds of times heavier than them (while large mammals can carry only a fraction of their weight) since the cross-section of their muscles is much larger in relation to the muscles' mass;
- as J.B.S. Haldane wrote, if a wet human has to carry a light film of water, a wet mouse has to carry its own weight in water, and a wet fly a mass of water many times heavier than its body;
- many small animals can climb on vertical surfaces or walk on the surface of water, since at their scale the surface forces (e.g. surface tension) are much stronger than volume forces (e.g. weight);
- insects can survive falls thousands of times higher than their body, while an elephant can fracture its bones by falling a few centimetres, since the force exerted by the fall depends only from mass and absolute height.
Mass, gravity and proportions[]
The most important stress that influences biomechanical structures is the pull of gravity. For example, the maximum height of an organism is given by the work of its structure to oppose its weight: E=mgh → h=E/mg. Since E (the energy output) and m (the mass) are both proportional to the volume, they cancel each other out, and therefore the height is only inversely proportional to gravity: h ~ 1/g. That means that, if the gravity doubles, the maximum height of an organism in divided in half. The same is true for the maximum height reached by jumping: since it's independent from mass, a jerboa and a kangaroo jump roughly to the same height, but if the gravity increases n times, this height will become 1/n times greater (that is, smaller).
Another consequence of changing gravity is the different burden upon the bones (or equivalent structures). The pressure the bones have to withstand is independent from the cross-sectional area, but it's directly proportional to weight, which is itself the product of mass and gravity: therefore, everything else being equal, the cross-sectional area of bones is directly proportional to gravity, and their radius to its square root (A ~ g, r ~ √g). If either the mass or the gravity double, the bones have to become √2 = 1.4 times wider.
Similar changes in the body shape can be computed in a similar way (remember that increasing the gravity n times is functionally equivalent to increasing volume by n times, and thus to increasing length by n3. For example, the torso width, neck length and leg length are all proportional to the square root of body length (~ √l).
What are the lower and upper limits of size? The smallest known organism capable of metabolic activity (therefore excluding viruses) is the parasitic bacterium Mycoplasma genitalium, with a diameter of 200-300 nm and a mass of about 10-13 kg; the smallest organism able to survive on its own (therefore excluding parasites too) in Pelagibacter ubique, about 400 nm long. The smallest known eukaryote, that needs a cell much more complex that any bacterium, is the alga Ostreococcus, 800 nm wide. Finally, the smallest known animal is the crustacean Stygotantulus stocki, 0.094 mm long. Being eukaryotes, animals need cells much larger than the minimum size; it has been calculated that a human being built with Mycoplasma-sized cells would have a mass of 50 mg and a height of 5 mm, though it's unlikely that cells that small would be able to support complex life.
As for the upper limit, largest sequoias, already built with a pillar-like shape, can weigh over 1000 tons, but they get to this size only because they don't have to move, something that puts much more stress on the structure; the largest known animals (blue whales and largest sauropods) have a mass of roughly 100 metric tons, or 105 kg. Perhaps on planets with weaker gravity the maximum mass would be higher: since mass is proportional to the cube of linear dimensions (such as height), and therefore to the cube of 1/g, we can deduce that the maximum mass is inversely proportional to the cube of gravity (m ~ 1/g3).
The limits of relative gravity on inhabitable planets are believed to be 0.2 and 2.2: that would lead leading to maximum masses of 12500 tons and 9.4 tons, respectively. Anyway, since the support that whales get from buoyancy doesn't seem to affect the result much, it's likely that the limit to size is given by other factors, such as the retention of metabolic heat and the increasingly difficult blood circulation.
| 0.2 g | 0.6 g | 1 g | 1.5 g | 2.2 g | |
|---|---|---|---|---|---|
| Height | 900 cm | 300 cm | 180 cm | 120 cm | 82 cm |
| Femur diameter | 1.6 cm | 2.7 cm | 3.5 cm | 4.3 cm | 5.2 cm |
| Mass | 10 tons | 370 kg | 80 kg | 24 kg | 7.5 kg |
| 0.2 g | 0.6 g | 1 g | 1.5 g | 2.2 g | |
|---|---|---|---|---|---|
| Height | 50 m | 17 m | 10 m | 6.7 m | 4.5 m |
| Femur diameter | 8.9 cm | 15 cm | 20 cm | 24 cm | 30 cm |
| Mass | 12500 tons | 460 tons | 100 tons | 30 tons | 9.4 tons |
Colonial organisms[]
Usually, a colony is simply a group of organisms of the same species that live close to each other; a true colonial organism is a combination of joined cells that are still able to survive on their own: a likely evolutionary precursor of multicellular organisms. Volvox is an example of such a colony (called a coenobium, since it has a fixed number of unspecialized cells) composed by 500-50000 flagellate cells, hold together by a gelatinous matrix of glycoproteins. A biofilm is a group of microorganisms that adheres to a surface through a slimy mass of polysaccharides or other chemicals: it's an important source of food for many detritivorous organisms.

The Portuguese man-o'-war (Physalia physalis).
There are examples of colonial organisms whose parts are in turn multicellular: they're called zoa (singular zoon) and the constituent individuals are called zooids. Zooids can be linked by soft tissue (as in corals and siphonophors) or share a common exoskeleton (as in pterobranchs and bryozoans). In some cases, mostly among siphonophors, of high specialization between zooids: for example, the "Portuguese man-o'-war" is composed by a pneumatophore (a translucid sac, 9-30 cm long, that floats on the surface of the sea), dactylozooids (tendrils up to 50 metres long, covered in stinging cells), gastrozooids (that digest killed preys) and gonozooids (that produce the sexual cells). This kind of association is rare on Earth, but on other planets it might be as common as simple multicellularity is here.
Structure and protection[]
Both static and mobile organisms will need rigid structures that oppose gravity allowing them to stand, and perhaps to move: the heaviest soft-bodied organism ever measured was a colossal squid (Mesonychoteuthis hamiltoni) weighing 500 kg, two hundreds times smaller than the 100 tons-limit to size given above, and without the support of water its maximum size would be even less. Boneless life has the advantage of flexibility - large octopuses can pass through holes a few centimeters wide (video), but a skeleton allows stability, protection and more efficient movement.
Early skeletons[]
The simplest form of skeleton is the hydrostatic skeleton, a cavity (coelom) or a system of cavities filled with liquid under pressure. Jellyfish, earthworms and starfish have such a system, which also works as muscle, providing both support and movement. Many animals have boneless mobile organs entirely composed of muscle (muscular hydrostats), such as the foot of mollusks (that becomes tentacles in cephalopods), the tube feet of echinoderms, the tongue of vertebrates and the trunk of elephants.
Sponges have a rudimental skeleton composed by spicules, small mineral elements with 2-5 pointed extremities sprinkled in the tissue; they're made of calcite in calcareous sponges, silica in glass sponges and spongin (a protein) or both spongine and silica in demosponges.
Echinoderms have a dermaskeleton, a simple rigid structure made up by calcite and magnesium oxide (MgO) plates, linked by the underlying and overlying layers of skin. A porous plate of madreporite filters seawater, which fills a net of canals, the water vascular system, with the triple function of locomotion, respiration and waste expulsion.
Biomineralization[]
Biomineralization is the process in which minerals are absorbed in the matrix of connective tissue to harden or stiffen it. Mineralized tissues include bone, horn, the enamel of teeth, shells and many others; their hollow structure make them 1000-10 000 stronger than the pure minerals. Earth lifeforms use:
- Calcite is the most stable mineral form of calcium carbonate (CaCO3). It's part of the shell of many planktonic organisms (coccolithophores and foraminifera, that form deposits of chalk and limestone), brachiopods and bryozoans, and partially of bivalve mollusks, of the spicules of calcareous sponges and of the skeleton of tabulate corals. The eyes of the trilobites, unique among the animals, had calcite lenses.
- Aragonite is another form of calcium carbonate (the third form, vaterite, is highly soluble in water, and it's therefore an unlikely replacement). It forms the shell of stony corals and all mollusks (though mixed with calcite in bivalves). Differently from calcite, it becomes unstable when magnesium becomes too rare in the environment relatively to calcium.
- Silica, or silicon dioxide (SiO2) forms the frustule (porous cells wall) of diatoms and some radiolarians and the spicules of glass sponges (see above). Many plants extract silica from the soil and store it in their tissues as phytoliths, which may have a structural function but mostly protect the leaves from grazing by wearing down the herbiores' teeth, especially in grass.
- Apatite is a group of phosphate minerals, which include hydroxylapatite (Ca10(PO4)6OH2), fluorapatite (Ca10(PO4)6F2) and chlorapatite (Ca10(PO4)6Cl2). Depending on its concentration in the mineralized tissue, hydroxylapatite forms bone (50%, mixed with proteins), dentin (which is found in teeth and in fish scales, 70%) and tooth enamel (90%).
- Strontium sulfate is used as the skeleton of Acantharea protozoans.
- Metals: in a process similar to biomineralization, some spiders and other arthropods accumulate solitary metal ions (mostly iron, copper, zinc and manganese) in their fangs, resulting in fangs that become progressively more metallic as the spider ages. While not common, such an adaptation could provide a definite advantage to the organism. A recently discovered gastropod (Crysomallon squamiferum), living in hydrotermal vents, has its aragonite shell reinforced by metallic plates of iron sulphide, specifically greigite (Fe3S4) and pyrite (FeS2); even the foot is covered in thick iron scales.
Some tissues undergo a process analogue to biomineralization but with organis substances: tendon and cartilage contain proteins (collagen and elastin); another protein, keratin, forms hair, feathers, claws, nails, hooves, the scales of reptiles, the beak of birds and turtles, horn, the quills of porcupines and the armour of pangolins. Polysaccharides can be concentrated around each cell, building a cell wall with cellulose (and sometimes the complex lignin) in plants and chitin in fungi.
The direct use of pure metals (as opposed to naturally occurring compounds such as oxides, sulfides, etc.) is very unlikely for different reasons; see this discussion on the forum.
External skeletons[]
There are two ways of building a complex skeleton: the exoskeleton surrounds the soft tissues, while the endoskeleton is surrounded by them. The jointed exoskeleton is the hallmark of arthropods
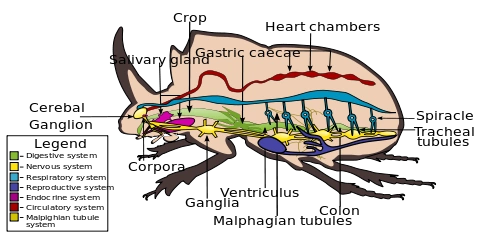
Internal morphology of a beetle (order Coleoptera). Notice the thick exoskeleton surrounding the body.
, in which case it's mostly made up of chitin, a polysaccharide, and permated by calcite: in insects it contains almost only chitin, while in trilobites, myriapods and crustaceans up to 40% of the exoskeleton is calcite. These substances form flexible jointed plates that cover all the body surface.
It has internal projections, apodemes, on which muscles are hinged; being entirely composed of chitins, apodemes are six times stronger and twice as stiff as vertebrates' tendons. This kind of skeleton gives to muscles a much larger area available for attachment, and it's much more resistant to bending, but it has two issues which worsen with size: first, while it's true that a hollow beam is stronger than a solid one when sustaining static loads, it's much weaker when it comes to dynamic loading, that is, when walking or falling, if the animal's mass isn't very little; second, it hinders the animal's growth. This last issues has been solved by arthropods with periodic moulting, though extraterrestrial insects in which new tissues expands the same plates, or different layers of exoskeleton are alternately redeposited one around the other, can be imagined. Anyway, no terran arthropod, both on land and in water, has ever grown beyond 2-3 metres long.
Less elaborate exoskeletons are found in other animals: brachiopods, bryozoans and some polychaete worms have one made up by calcite; mollusks have a distinctive dorsal tissue, the mantle or pallium, which secretes calcium carbonate and conchiolin (a protein). It's a very hard shell, having a point of articulation only in bivalves, and in some brachiopods. Even some vertebrates have developed an exoskeleton: ostracoderm fish had a head armour of bone, cartilage and dentin, while turtles and armadillos have a bone carapace, and pangolins an armour composed by hair-derived keratin, which in other mammals also forms horns and quills.
Internal skeletons[]


Collection of skeletons and skulls from different vertebrate species.
While the spicules of sponges, the echinoderm's dermaskeleton and the spongy aragonite cuttlebone of cuttlefish can be considered internal skeletons, a complex jointed endoskeleton is typical of chordates, in which is centered around a dorsal spinal cord, often a skull that protects the brain, a ribcage around the thoracic organs and protrusions that stiffen the limbs. This skeleton is composed either by cartilage (matrix or proteins, mostly collagen and elastin) or bone (ossein and calcium phosphate) with cartilage elements.
Since bone can resist to a compression up to 1700 atm (that is, a weight equivalent to 1 kg on each square cm), that means that an animal with a mass of 5000 kg (assuming a gravity equal to Earth's) needs, at the very minimum, a total leg cross-section of 5000/1700=2.9 square centimetres, or, assuming one cylindrical leg, a bone diameter of 1.9 cm, while a cylindrical hardwood trunk (strength of 500 atm) should be 3.6 cm wide, a cartilage foot (strength of 350 atm) 4.2 cm wide, and a muscle foot (strength of 10 atm) 25 cm wide.
Clearly, Earth's animals have much larger legs than this: that's because they're built not just for standing, but also for walking, running and leaping. When humans run, the foot strikes the ground with a force about 2.5 times larger than the body weight (the total force is therefore 3.5 times the weight), and in animals with less anatomical specialization for running it might be even higher. Putting all of this together, the formula to get the minimum legbone diameter is d = 2√(mgF/πnC), where m is the total body mass, g the relative gravity of the planet, F the magnitude of the total force relative to the body weight when running (we already got 3.5 for humans), n the number of legs and C the compressive strength of the bone. Let's assume 5000 kg of body mass again, Earth's gravity, F=4, six legs and true bone (1700 atm): we obtain thus legbones 2√(20000/32000) = 2√0.63 = 1.6 cm wide, for a total cross-section area of 8.0 square centimetres.
An interesting variation of the endoskeleton is found in the basket skeleton (see halfway down here), a sort of "wicker basket" of interconnected bones that surrounds the soft body interior, similar to an enlarged version of the see urchin dermaskeleton or the hollow bones of birds; such a structure would combine great strength and lightness. Other alternatives proposed in Xenology include multiple spinal cords, helical ("corkscrew") arrangement of internal fibres, telescoping bones (one retracting inside another), etc.
Movement[]
Muscle is a tissue composed by protein filaments; when they receive an electrical signal, they contract, shortening the muscle fibres and thus exterting a force. In arthropods, muscle are anchored on the internal surface of the exoskeleton, while in chordates they're tied to bones by tendons. Skeletal muscles are classified in four categories according to the arrangement of fibres:
- In parallel muscles (e.g., the biceps brachii), fibres are all parallel to the longitudinal axis of the muscle. They can be flat or spindle-shaped, with tendons at both ends; they can contract up to two thirds of their maximum length, exerting a force of 36 N per square centimetre (3.6 atm).
- In convergent muscles (e.g., the pectoralis major), fibres are spread from the attachment point, where a tendon or a similar structure is located. They're broad, flat and fan-shaped, and not as strong as parallel muscles, but they can pull in several different directions.
- In pennate muscles (e.g., the extensor digitorum, unipennate, and the rectus femoris, bipennate), fibres form the same angle with the longitudinal axis (they're parallel to each other, but not to the tendon). They contain more fibres per unit of volume, and they're therefore stronger than other muscles.
- In circular muscles or sphincters (e.g., the orbicularis oris), fibres are arranged in circle around a central aperture, restricting the diameter when they contract. They are often located around orifices and passageways to regulate the flow of matter (or light).
Given the physics of levers, the distance of the muscle insertion point on a bone (wheter it is on the inside or on the outside) from the articulation, for example that between the insertion of biceps on the radius and the elbow, influences the strength and speed of the flection: if the insertion is closer to the articulation, it makes the movement faster; if it's farther, it makes it stronger. Since muscle fibres can only pull, every bone needs at least two muscles on opposing sides, such as biceps and triceps for the forearm.
A variation of this is found on Snaiad, whose "vertebrates" have hydraulic muscles derived from a vascular system similar to that of Earth's echinoderms: they're fluid-filled sacs with a partitioning that makes them resistant to puncture, which push the bones instead of pulling them. This has consequences on the morphology of limbs. This system is not entirely unheard-of: jumping spider stiffen their legs before a jump by building up blood pressure. In a similar way, Epona's pentapods use a system of fibres ("myoskeleton") that extend when they absorb water by osmosis. Myoskeletal rods can also bend (by extending only fibres on one side) or twist (extending fibres in a spiral pattern); this system works both as musculature and skeleton.
Movement in response to an environmental stimulus, without mediation by the nervous system, is called tropism. Depending from the type of stimulus (see here how organisms can perceive them), tropism can be:
- Chemotropism (chemicals), such as the movement of roots towards useful minerals and away from toxins.
- Hydrotropism (water), a particular form of chemotropism where roots "search" water.
- Heliotropism or phototropism (light), such as plants growing towards light sources or turning to face the Sun; it can be achieved by auxin, a hormone that's destroyed by light and stimulates tissue growth, therefore elongin the darker side of a stem to curve it.
- Geotropism or gravitropism (gravity), such as the growth of plants and fungi with the stem upwards regardless of the ground inclination.
- Thermotropism (temperature), such as the curling of Rhododendron leaves in response to cold.
- Electrotropism (electric fields), such as the particular growth, still not entirely understood, of pollen tubes under electrical stimulation.
- Thigmotropism (touch or contact), such as the coiling of vine tendrils over supporting objects, the leaves of Mimosa pudica folding when touched or those of the Venus flytrap closing over trapped insects.
Locomotion[]
Also see here and following pages Most lifeforms have to move to survive, be they microscopic algae and bacteria, burrowing parasites, swift hunters or giant filter-feeders - with some exceptions, such as fungi, most plants and some animals, that live anchored to the ground, and are called sessile. Other organisms have instead developed a huge variety of means of locomotion, which can be grouped in two kinds: movement inside a fluid and on a solid ground.
See this discussion on the forum about manipulators (portions of limbs or other structures used to manipulate tools and other objects).
Locomotion in a fluid medium[]
When a body is immersed in a fluid in relative motion (that is, both when the body is motionless and the fluid moves and when the opposite occurs), it experiences four forces that dictate its movement:
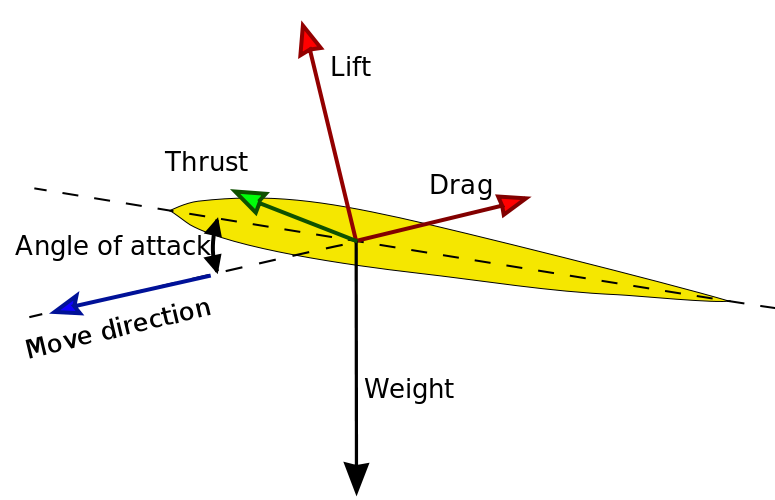
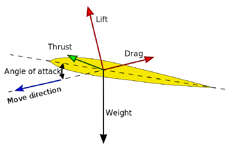
Aerodynamic/hydrodynamic forces acting on an aerofoil/hydrofoil.
- Weight, the product of body mass and gravity; it alwasy points towards the centre of the planet. If the fluid's density is significant compared to that of the body, the weight can be partially countered by buoyancy (see "floating" below).
- Thrust, the force exerted by the body's propulsion, as a reaction to the fluid pushed backwards.
- Drag, the force that opposes the movement (it has thus the same direction, but opposite verse), given by the friction of the body surface with the fluid.
- Lift, the force perpendicular to the movement. If the aerofoil/hydrofoil moving through the fluid, at a certain angle of attack, deflects it with more force above than below, the fluid exerts a reaction force that pushes the body upward. Lift is proportional to the fluid's density, to the surface of the aerofoil/hydrofoil and to the square of the speed of movement.
Liquids and gases act in very similar ways: swimming and flight are basically regulated by the same physics, taking into account the far lesser density of air. A body with a thin cross-section that produces strong aero/hydrodynamic forces (such as fins, wings, blades, sails) is called aerofoil (in air) or hydrofoil (in water).
Swimming[]
The Froude's Law states that the maximum velocity of an organism swimming in water has to be proportional to the square root of the ratio between muscular work and water resistance (themselves proportional to the volume and to the surface area of the body), which is the linear length: therefore, v ~ √l.
However, the resistance the fluid exerts on the swimming animal also depends from its viscosity: if we include this in the formula, we find that the maximum velocity in liquids other than water is also inversely proportional to the square root of the viscosity. That is, an animal swimming in hydrogen cyanide will be 1/(√0.21) = 1/0.46 = 2.2 times as fast as the same animal swimming in water. Nearly all the most likely thalassogen are far less viscous than water, and thus require less energy to swim in them, with the exception of molten sulfur.
| Thalassogen | Density | Viscosity | Thalassogen | Density | Viscosity |
|---|---|---|---|---|---|
| Helium | 141 g/L | 0 | Cyanogen | 950 g/L | n/a |
| Hydrogen cyanide | 397 g/L | 0.21 | Water | 1000 g/L | 1 |
| Methane | 424 g/L | 0.012 | Carbon disulfide | 1266 g/L | 0.40 |
| Ammonia | 683 g/L | 0.29 | Nitrogen dioxide | 1378 g/L | 0.48 |
| Carbon monoxide | 789 g/L | ~0.067 | Sulfur dioxide | 1464 g/L | 0.013 |
| Hydrogen sulfide | 950 g/L | 0.33 | Sulfur | 1800 g/L | 94 |
Helium is at superfluid temperatures.
As for the means of propulsion:
- Undulation: the most simple way to advance in water is moving sideways the body, creating undulations that push water backwards and, by reaction, the body forward. Lampreys, eels, sea snakes, mosasaurs and many other animals swim this way; fish, sharks and ichthyosaurs move sideways only a tail equipped with large fins, while seals, otters and whales flex their body up and down. Fins or flukes perpendicular to the direction of the undulation (therefore vertical in fish and horizontal in whales) help to push water with more strength. As a variation, rays, cuttlefish and anomalocaridids swim by rippling lateral membranes, keeping the body still (also see here, with different patterns of undulations).

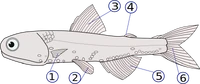
1: pectoral fins (paired); 2: pelvic fins (paired); 3: dorsal fin; 4: adipose fin (only in some species); 5: anal fin; 6: caudal fin
- Propulsion through lift: In many vertebrates (e.g. sharks, penguins, sea lions), long and wide horizontal fins act as hydrofoils, create lift; in flatfish, the whole body becomes the hydrofoil. These organisms need to reduce the water resistance, through a tapered spindle-shaped body and a smooth surface; grooves and tubercles, though, can reduce drag by breaking the flow of water in turbulent channels, as they do on a humpback whale's flippers.
- Propulsion through drag: In a similar way, other organisms (e.g. sea turtles, plesiosaurs) use fins or other appendages as oars, exploiting drag rather than lift, pushing water backwards as shrimps and swimming crabs do with pleopods. Birds that row with their feet, such as ducks and grebes, have legs placed at the rear end of the body, which impairs their land locomotion. These organisms also need a streamlined body and smooth skin: sea turtles even have a lower shell than land tortoises, while duck feathers and otter fur are covered in an oily substance over which water flows with little unwanted drag.
Fins can have different functions depending from their position on the body. Fins aligned to the anteroposterior axis, such as the dorsal and anal fin, make the animal stable, preventing it to fall on its side; paired fins, such as the pectoral and pelvic fins, provide lift (see below) and allow to change direction. - Jet or reaction propulsion: cephalopods (cuttlefish, octopuses, squids) and jellyfish swim by drawing water in a body cavity and expel it with a strong muscular contraction, moving by reaction in the opposite direction relatively to the jet of water; scallops swim by "clapping" the shells, which quickly expels water from all sides, while the flat shape acts as a hydrofoil, preventing it from sinking. This method of locomotion is more efficient when the fluid is ejected through a small opening. In a variation, salpae and Furahan tubesharks swim by passing water through all the body, pushing it backwards with a sequence of muscular contraction called peristalsis (also see here).
- Osmosis: Xenology proposes an exotic form of locomotion in which an animal, swimming in freshwater or otherwise a very diluted liquid, exposes a frontal semipermeable sac filled with highly concentrated salts. The osmotic pressure forces water in the sac to dilute the salts; this water is pumped out in a rear sac, from where it can be expelled as in jet propulsion. Pure water exterts on seawater a pressure of 25 atm, or 250 newtons of force on each exposed square centimetre. This method could evolve in oceans containing poisonous salts to be kept out of the body, and/or too viscous for other methods to be convenient (such as molten sulfur oceans).
- Floating: It's possible to float on the sea surface by developing light tissues or gas-filled sacs so that the overall body density is lower than the density of the thalassogen. The force exerted upwards by the sac (or equivalent structure) F = V·(dt-ds)·g, where V is the sac volume, dt the thalassogen's density, ds the sac's density and g gravity; besides itself, the sac can sustain a mass of m = V·(dt-ds). If the mass grows above this value, the organism sinks; if it falls below, it rises to the surface; if it's equal, the organism can remain stable at any depth without effort. Gravity doesn't affect the result, since it has the same effect both on upward and downward forces. A floating organism could then move itself with jets of water, paddle-like feet rowing in sequence, structures similar to paddlewheels or even sails - as those of the hydrozoan Velella and the ocean phantom - perhaps guiding its direction with a rudder-like appendage.
Even organisms that don't actually float have adaptation to counteract their weight: siphonophores, many cephalopods and teleost are equipped with bladders full of gas, such as the swim bladder (full of air, sometimes with more oxygen); cetaceans have thick layers of fat, which is lighter than water; many sharks have asymmetrical tailfins longer in the ventral side than in the dorsal one, to push themselves upwards when swimming. Kelp, a large seaweed, keeps itself straight with pneumatocysts full of different gases.
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
.png){kind=link}
| Dwarf seahorse (Hippocampus zosterae) | 0.15 km/h | 0.83 l/s | Drag |
| Green sea turtle (Chelonia mydas) | 3 km/h | 0.74 l/s | Drag |
| American eel (Anguilla rostrata) | 3.9 km/h | 1.1 l/s | Undul. |
| Emperor penguin (Aptenodytes forsteri) | 7.5 km/h | 1.7 l/s | Lift |
| Sea otter (Enhydra lutris) | 9 km/h | 1.8 l/s | Undul. |
| California sea lion (Zalophus californianus) | 22 km/h | 2.7 l/s | Lift |
| Bottlenose dolphin (Tursiops truncatus) | 35 km/h | 3.2 l/s | Lift |
| Gentoo penguin (Pygoscelis papua) | 36 km/h | 14 l/s | Lift |
| Common octopus (Octopus vulgaris) | 40 km/h | 15 l/s | Jet |
| Great white shark (Carcharodon carcharias) | 40 km/h | 2.3 l/s | Lift |
| Blue whale (Balaenoptera musculus) | 50 km/h | 0.46 l/s | Lift |
| Flying fish (Cypselurus californicus) | 57 km/h | 32 l/s | Lift |
| Bluefin tuna (Thunnus orientalis) | 70 km/h | 9.7 l/s | LIft |
| Swordfish (Xiphias gladius) | 97 km/h | 13 l/s | Lift |
| Sailfish (Istiophorus sp.) | 110 km/h | 10 l/s | Lift |
Note: speed is given both in kilometres per hour and in body lengths covered per second.
Flight[]
There are several different degrees of flight specialization. Since no living tissue has a density even just comparable with air (though some biologically produced gases could), and buoyancy is thus usually negligible, most forms of aerial locomotion require active movement.
| Gas | Density | Viscosity (air=1) | Gas | Density | Viscosity (air=1) |
|---|---|---|---|---|---|
| Hydrogen | 0.0899 g/L | 0.47 | Oxygen | 1.43 g/L | 1.1 |
| Helium | 0.179 g/L | 1.1 | Argon | 1.78 g/L | 1.2 |
| Water vapour | 0.596 g/L | 4.7 | Nitrous oxide | 1.80 g/L | 0.79 |
| Methane | 0.667 g/L | 0.58 | Carbon dioxide | 1.84 g/L | n/a |
| Ammonia | 0.696 g/L | 0.53 | Ozone | 1.96 g/L | n/a |
| Neon | 0.900 g/L | 1.7 | Sulfur dioxide | 2.62 g/L | 0.68 |
| Carbon monoxide | 1.15 g/L | 0.95 | Sulfur trioxide | 3.12 g/L | n/a |
| Nitric oxide | 1.23 g/L | 1.0 | Krypton | 3.75 g/L | 1.3 |
| Nitrogen | 1.25 g/L | 0.95 | Xenon | 5.90 g/L | 1.2 |
| Hydrogen sulfide | 1.39 g/L | 0.63 |
Note: the density of a mixture of gas can be derived from those of its components. For example, Earth's air is 78% nitrogen and 21% oxygen: thus, its overall density is (0.78*1.25)+(0.21*1.43) = 1.28 g/L. Note 2: the density of gases is directly proportional to atmospheric pressure: thus, Earth's air at 3 atm would have a density of 1.28*3 = 3.83 g/L. Heating a gas makes it less dense, but the change is usually negligible.
Some mathematical relationships: speed (especially stall speed, the minimum speed that still allows the animal to stay in air) is inversely proportional to the square root of the wing surface (v ~ 1/√S) and also to the square root of the body length (v ~ 1/√l), so, given identical conditions, an animal with wings four times smaller than another can (and has to) fly twice as fast; it's also proportional to the square root of the atmospheric pressure (v ~ √P) and of gravity (v ~ √g), so an animal can fly half as fast on a planet with a pressure of 4 atm or a gravity of 4 g. Xenology gives a formula to estimate wing surface (in square metres), being S = 0.24(mg)0.82/P, where m is the animal mass (in kg), g the relative gravity and P the pressure (in atm). See here for information about the origin and physics of bird flight.
- Parachuting: The simplest form of aerial movement, essentially a controlled fall. Many spiders, especially young ones, allow the wind to carry them suspended from a strand of silk, often reaching heights of several kilometres and venturing over seas; dandelion seeds do the same. In planets with strong winds and a thick atmosphere, such a method would allow to travel hundreds of kilometres per day. Parachuting and gliding are very similar: but while parachuting organisms use their aerofoils for drag and usually fall vertically more than they travel horizontally, gliding ones privilege lift and horizontal movement.
- Gliding: Simple membranes to generate lift and glide have evolved at least six times in mammals (sugar gliders, the greater glider, anomalures, true flying squirrels, colugos and Volaticotherium), ten times in reptiles (flying dragons, flying snakes, Holaspis lizards, four species of geckos, three extinct groups), twice in amphibians (Rhacophoridae and Hylidae flying frogs), three times in fish (freshwater butterflyfish and hatchetfish, true flying fish), four times in insects (gliding ants of three subfamilies and gliding bristletails) and once in mollusks (flying squids).
Differently from powered flight, gliding does not involve thrust: the speed that produces lift is given only by gravity. Gliding organisms rarely have true wings (with exceptions such as samarae, a kind of fruit, and gliding ants), and their L/D ratio (or "lift-to-drag ratio") is relatively low. - Flapping: It's the most energy-consuming method of flight, involving a cyclic movement of wings to generate thrust, increasing speed and therefore lift. Flapping flight has evolved four times on Earth, in insects, pterosaurs, birds and bats. While vertebrate wings have articulations and can be folded to reduce drag, insect wings remain rigid during flight: they're moved by muscles inserted at their base (in mayflies and dragonflies) or that deform the whole body (in other insects); wings are flipped between downstroke and upstroke. Bats, instead, flap only their digits. In every case, the downstroke needs to be stronger than the upstroke.
Another relevant factor is wing loading, the ratio between the bird mass (though gravity should be taken into consideration) and wing area.
| Swallow | 0.15 kg/cm2 |
| Passerines | 0.2-0.4 kg/cm2 |
| Hawks | 0.3-0.5 kg/cm2 |
| Waterfowl | 0.8-1.0 kg/cm2 |
| Grebe | 1.2 kg/cm2 |
| Loon | 1.4 kg/cm2 |
| Vulture, condor | 1-3 | Chickadee | 27 |
| Hawks and eagles | 2-3 | Hummingbird | 80-90 |
| Pigeon | 5-15 | Hornet | 100 |
| Cabbage butterfly | 12 | Housefly | 190 |
| Mockingbird | 14 | Honeybee | 250 |
| Nectar-feeding bats | 15-17 | Mosquito | 500-600 |
| Desert locust | 17-20 | Gnat | 1000 |
| Flying squirrel | 2 |
| House sparrow | 4 |
| Herring gull | 10 |
| Albatross | 20 |
- Soaring: While able to flap their wings, many birds, especially of large size and low loading (and probably largest pterosaurs), developed adaptation to soaring flight, namely long, narrow and pointed wings (see below), stiffened by tendons in albatrosses. This allows them to fly for long distances (even thousands of kilometres) without flapping, particularly on oceans, which offer no resting place. Soaring animals can extract additional lift from phenomena such as thermals (columns of warm and rising air), ridge lift (wind blowing upwards on a slope), convergence zones (warm inland air pushed upwards by colder sea air), etc.
- Hovering: This form of flight allows the animal to remain still in air, thanks to wing flaps that exert the same force with upstroke and downstroke. Since there's no movement through air, all the lift has to be generated by flapping; hovering flight requires therefore an extremely fast flapping (at least several tens of beats per second) and is more suited to small organisms, where muscle strength overcomes weight. Hovering is practiced by most insects, but among vertebrates it's confined to some birds (mostly hummingbirds) and nectar-feeding bats; the largest hovering vertebrate, the pied kingfisher, weighs only 70-90 grams.
- Ballooning: A type of flight unknown on Earth requires the presence, in sacs or cavities, of a gas or mixture of gases less dense than air (given Earth's atmosphere and biochemistry, likely possibilities would be hydrogen, methane and carbon monoxide). Gravity has no effect on ballooning, but the size of the needed sac depends from the difference of density between the gases: a litre of hydrogen can sustain up to (1.28 g/L-0.0899 g/L) = 1.19 grams, a litre of carbon monoxide (1.28-1.15) = only 0.13 grams. Therefore, a body 10 kg heavy needs a balloon of hydrogen of (10000/1.19) = 8400 L = 8.4 m3 , or a ballon of carbon monoxide of (10000/0.13) = 77000 L = 77 m3. Ballooning organisms will tend thus to become very big but with denser structures tripped to the bone, except on planets with an extremely high atmospheric pressure (with Venus' pressure, the 10 kg animal would need just 92 litres of hydrogen).
Furaha's associated blog contains an extensive tractation of ballooning organisms (which are called "ballonts"): since the mass of the sac casing increases proportionally to the surface and the efficacy of the gas to the volume, ballooning doesn't work on small scale[2]; it could evolve perhaps from floating animals, which could start small, grow and then take the air. Interesting examples of ballooning organisms include Furahan zeppeloon, Blue Moon balloon plants and Darwin IV eosapien.
While the number of wings (aerofoils) doesn't affect the result, as long as there is a certain overall wing surface available, most insects have two pairs of wings, one of which often specialized for another functions: a protective shield (elytra) in beetles, gyroscopes to control flight (halteres) in flies and mosquitoes; there is a Furahan flyer which uses a pair of mobile wings for thrust and a pair of rigid wings for lift. Every other flying animal on Earth has no more than one pair of wings, though more pairs could be use for different kinds of flight according to their shape. In fact, different regular wing shapes have been identified in birds, with different functions:
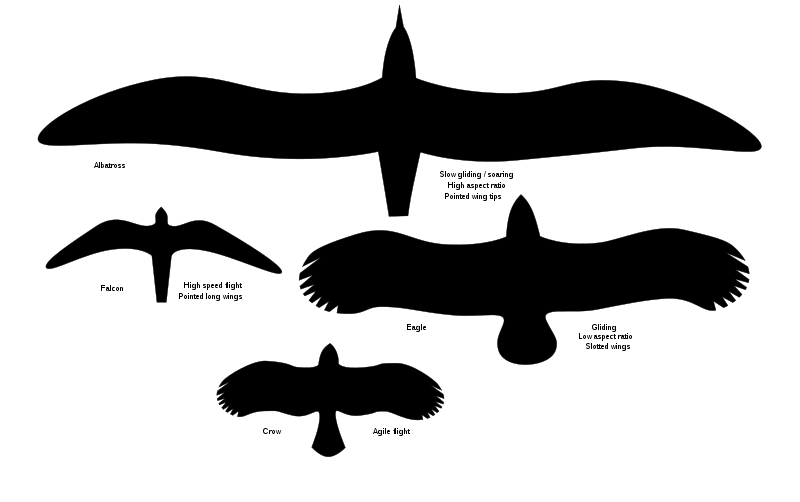
{kind=link}
Comparison of wing shape in a crow, an eagle, a falcon and an albatross.
- Elliptical wings. Birds that live in covered environments (e.g. woodland), such as pheasants and grouses, have short, broad and slightly arched wings that allow a quick take off and high manoeuvrability. Wing loading is high.
- High speed wings. Birds that pursue preys, such as swallows, swifts and falcons, have thin, tapered wings, triangular and pointed backwards, that allow an extremely fast flight (up to hundreds of kilometres per hour).
- Soaring wings. Albatrosses, gulls and frigatebirds, which fly for thousads of kilometres on the ocean, have very long, narrow and tapered wings, with sharp points and compact primary feathers. Giant pterosaurs had probably wings similar to these. Wing loading is low.
- Slotted soaring wings. Inland soaring birds, such as condors, hawks and ravens, have long and broad wings (long about three times as they're wide), with slightly divergent primary feathers that work as separate airfoils; similarly for the slowly flapping herons and egrets whose wings are very large, broad and arched; storks and some vultures have yet similar wings with long primary feathers to make use of each air current.
- Hovering wings. Hummingbirds have pointed, tapered wings, which rotate at the shoulder rather than the wrist; this allows them to beat backward with the same force as forward, so that the movements cancel each other.


{kind=link}
A graph plotting birds according to the shape (aspect) and relative size (loading) of their wings. Based on a graph at http://people.eku.edu/ritchisong/554notes2.html.
| Housefly (Musca domestica) | 7.2 km/h | 200 l/s | (insect) |
| Dragonfly (Anax parthenope) | 29 km/h | 81 l/s | (insect) |
| Pigeon (Columba livia) | 59 km/h | 47 l/s | Elliptical |
| Pheasant (Phasianus colchicus) | 90 km/h | 83 l/s | Elliptical |
| California condor (Gymnogyps californianus) | 90 km/h | 20 l/s | Sl. soaring |
| Gannet (Morus bassanus) | 100 km/h | 29 l/s | Soaring |
| Eider duck (Somateria mollissima) | 113 km/h | 52 l/s | Elliptical |
| Horsefly (Hybomitra hinei) | 145 km/h | 1006 l/s | (insect) |
| Frigatebird (Fregata aquila) | 153 km/h | 57 l/s | Soaring |
| White-throated swift (Hirundapus caudacutus) | 170 km/h | 190 l/s | High speed |
| Red-tailed hawk (Buteo jamaicensis) | 195 km/h | 108 l/s | High speed |
| Peregrine falcon (Falco peregrinus) | 322 km/h | 203 l/s | High speed |
Locomotion on a surface[]
Regardless of the number of legs, modeling each step as an inverted pendulum with its fulcrum in the ground gives that the maximum speed of a walking (not running) animal is proportional to the square root of leg length and gravity (v ~ √(lg)), which means that an animal will walk twice as fast if its legs are four times longer or the gravity is four times stronger, with a margin of error of about ± 10%. On Earth, it's roughly v = 2.2√l, so an animal with a leg 1.5 m long will have a maximum walking speed of 2.2√1.5 = 2.2·1.2 = 2.7 m/s = 9.7 km/h.
Limbless land locomotion[]
- Undulation: The simplest method of propulsion on land is arguably, as in water, the undulation of the body. Snakes and other legless lizards (and eels, when they crawl out of water during migrations) bend their body sideways thanks to circular muscle rings, pushing against irregularities in the ground; the wave moves at the same speed of the snake, and thus each point of its body follows the same path of those preceding it. Vertical undulation, though, is almost useless on solid ground (in fact, it may be the reason there are no limbless mammals): the inchworm moves, very slowly, in a similar way, having two pair of legs at each extremity.
- Sidewinding: It's employed mostly on flat terrains without irregularities, such as mud flats, loose soil and sand dunes. The body is always static when in contact with the ground; the snake seems to "throw" the head forward, and then parts of the body follow, proceeding diagonally and forming peculiar straight or J-shaped parallel tracks, each of which as long as the snake's body.
- Concertina: It's typical of enclosed or otherwise limited spaces that do not allow sidewinding. Tunnel concertina involves pressing against the walls with the lateral bends of the body, while arboreal concertina bends are used to grip branches, and the body follows a fixed path as in lateral undulation. It's extremely slow, rarely faster than 0.05 body lengths per second. Leeches have a suction cup at each end of the body, which they use to anchor themselves to the ground.
- Rectilinear locomotion: Heavy-bodied snakes such as boas and pythons can advance is a straight line by lifting one section of the body at time. it's extremely slow but noiseless, and useful for stalking predators. Large ventral scales (scutes) pin to the ground, pushing on it backwards. Slugs, snails, limpets, some flatworms and even seals move foward by rippling their underside (pedal waves); penguins commonly slide on their abdomen down snow slopes.
- Peristalsis: Earthworms and other worms move with a series of muscular contractions that make alternate parts of the body wide and short or stretched and thin. Like snake scutes, they have tiny bristles (setae), pointed backward, that force each section of the body to stretch forward.
- Amoeboid locomotion: Some slime molds crawl on the ground by moving back and forth their gelatinous mass and extending tendrils, while some other form a moving slug-like body (grex). The amoeboid locomotion is very slow, usually less than 0.1 body lengths per second.
- Rolling: Organisms that actively roll their body in a loop and move by rolling include tiger beetle larvae, the Nannosquilla mantis shrimp, myriapods and salamanders; the caterpillar Pleurotya can roll at 40 cm/s, forty times its walking speed, the wheel spiders at 1 m/s (20 revolution per second); tumbleweeds detach from their roots and roll to disperse seeds. Other animals, most of them armoured (pangolins, hedgehogs, armadilloes, armadillo lizards, pebble toads, woodlice, trilobites) commonly assume a spherical shape and roll assisted by gravity or by their own propulsion.
- Wheels?: Despite the simplicity of a wheel design, no true rotating structure has ever been found in a macroscopic organism: the closest example is the bacterial flagellum, a protein tube which rotates in a ring driven by a flow of protons. When it comes to multicellular species, though, the need arise to grow the wheel from living cells, which in turn requires a flow of nutrients; since a wheel has to turn around an axle it's not possible to run nerves and blood vessels to it (unless it's mounted on an exceptionally elastic stalk and periodically unwound). The writer Philip Pullman devised a solution based on symbiosis: the Mulefa lock circular and perforated seed pods on two legs, propelling themselves with other two.
Moreover, wheels require a flat, even and rigid ground to be more effective than limbs, which are far more useful on irregular or soft ground. Natural approximations of roads could be found - ice, for example, or rock highly eroded by wind - but organisms that build road for themselves would be victims of strong parasitism, given that each builder would provide useful work for many other individuals. Finally, wheels are not much versatile, dealing poorly with vertical obstacles and not being easily adapted for tasks other than locomotion.
Leg structure and posture[]
In the animal kingdom, the number of limbs varies from none to many dozens, though the highest number show scarce versatility (centipedes and millipedes move all their legs in a wavelike sequence, unable to control them one at time). The most successful groups of limbed land animals, tetrapods and insects, typically have respectively four and six limbs; a higher number is unnecessary, but could evolve due to evolutionary contraints. Since a plane can be identified with three points, three legs is the minimum number of legs needed to keep the animal always on the ground without a complex balancing system. Bipeds usually have a horizontal stance where head and tail balance each other's weight (as in theropod dinosaurs and kangaroos); monopods, such as TFIW desert hopper, Leap Snouters or the Eponan Springcroc, don't have any other feet on the ground when they "walk", so they're forced to make only very short steps, especially in high gravity.
Since all land animals have a bilateral symmetry, the number of limbs is usually even; odd-numbered functional appendages tend to be highly specialized organs (wasp ovipositor, scorpion stinger) or grasping structures (elephant proboscis, prehensile tails); if they derive from fins, as it happened in tetrapods, they're likely to be paired, given the need for paired fins to assure stability when swimming. According to an evaluation on Furaha's blog, the compromise between weight and strength of the limbs means that larger animals need fewer legs; since they're likely to move slowly, and thus will need to keep three feet on the ground at all times, four legs seems to be the optimum number of limbs for the heaviest moving organisms.
The number of digits is also likely to be highly variable. The general tendence seems to be reduction: the first tetrapods had 6-8 digits that were soon reduced to 5, and since then no known tetrapod has ever had more than five digits (in some ichthyosaurs there was six, but only fused together in a paddle-like limb rather than differentiated). Such reduction is greater in running animals: ostriches, deer, gazelles, horses and litopternans have reduced their digits to two or even one. Soft muscular limbs (tentacles) are not good to move out of water, since they don't sustain weight well (though hydraulic muscles could produce better weight-bearing tentacles).
| Name | N. of limbs | Examples |
|---|---|---|
| Nullipeds | 0 | Snakes, eels, worms |
| Monopods | 1 | Snails; rotifers; some clams; seahorses |
| Bipeds | 2 | Kangaroos; jerboas; sealions; many dinosaurs and all birds; basilisk; sea cucumber larvae; lemurs (on the ground); humans |
| Tripeds | 3 | Kangaroos (when moving with the tail); tripodfish |
| Quadrupeds | 4 | Most amphibians, reptiles and mammals; inchworms; praying mantises; water scorpions; humans (with hands) |
| Pentapods | 5 | Almost all sea stars; New World monkeys and many tree lizards (with the prehensile tail); elephants (with the proboscis) |
| Hexapods | 6 | Almost all insects |
| Heptapods | 7 | Springtails (with the furcula); trilobite beetle (with the abdomen); ichneumon wasps (with the ovipositor) |
| Octapeds | 8 | Octopuses; shrimps (walking legs only); spiders, scorpions, mites; some sea spiders |
| Nonapeds | 9 | Scorpions (with the stinger) |
| Decapods | 10 | Prawns and lobsters (walking legs only); crabs and scorpions (with claws); horseshoe crab; squids; some sea spiders |
| Multipeds | Various |
12: Some sea spiders; many fly larvae and caterpillars; 14: Prawns and lobsters (all legs); many caterpillars; 16: Many caterpillars; 18: Pauropods; sawfly larvae; 22: Shrimps (with all appendages); 24: Symphylans; ~30-40: Most centipedes (Lithobius, Scutigera, Scolopendra); ~60-90: Nautilus; ~100: Trilobites; ~200: Centipede Orya barbarica; ~64-400: Most millipedes; 600-750: Millipede Illacme plenipes |
| Name | N. of digits | Examples |
| 0 | Birds (wings); squids and octopuses | |
| Monodactyls | 1 | Horses; Thoatherium (extinct); titanosaurs (extinct); crabs; spiders |
| Didactyls | 2 | Camels, sheep, pigs, deer, giraffes, antelopes, pronghorn; ostriches; two-toed sloth; pig-footed bandicoot (extinct); flies |
| Tridactyls | 3 | Most ratites; bandicoots; rhinoceros; early horses, litopternans, chalicotheres |
| Tetradactyls | 4 | Colobi and spider monkeys; hyenas and dogs; tapirs; hippos, armadilloes; elephant shrews; anisodacyls (3 toes forward, 1 backward): birds of prey, songbirds, chickens, most theropods; zygodactyls (2 toes forward, 2 backward): woodpeckers, parrots, cuckoos, owls, chameleons; pamprodactyls (4 toes forward): swifts |
| Pentadactyls | 5 | Most primates, carnivorans and reptiles; elephants; dolphins and walruses (vestigials) |
| Multidactyls | Various | 6: Giant panda; 6-8: Basal tetrapodes ("ichthyostegalians"); 10: frogfish |
As for the shape of limbs, the speed of movement directly influences their length. Given the relationship between speed and leg length, faster animals will extend their legs walking only on the very tip (digitigrades), or even on the nails (unguligrades), while in those who need less speed but more weight-bearing the last segment of the leg (the foot) will entirely touch the ground (plantigrades). Wide feet are also useful on shifting grounds such as mud and sand, while animals moving on rocks will be mostly unguligrades (with the extreme case of the klipspringer, an african antelope which stands on vertical hooves). Species with long or fragile digits (the grasping fingers of gorillas and chimpanzees, or the curved claws of anteaters and ground sloths) walk on their knuckles.
Posture (the way the body is supported by legs) can be roughly divided in sprawling (legs sticking out sideway) and erect (legs vertical under the body). Sprawling posture is used by amphibians and lizards; erect posture by dinosaurs, birds, mammals; an intermediate (semi-erect) posture is used by insects, monotremes and larger reptiles, such as crocodiles and monitor lizards. The distance of articulations from the body with a sprawling posture creates a torque which makes the body weight much more onerous to support; thus, the sprawling posture is not adapt to large-sized animals (especially under high gravity), and legs with many jointed segments are best folded under the body. Sprawling legs, though, can be useful to resist to lateral forces, such as water currents, tide reflux and (on very small scales) wind, or perhaps when moving on vertical surfaces.
Gaits[]
Also see here
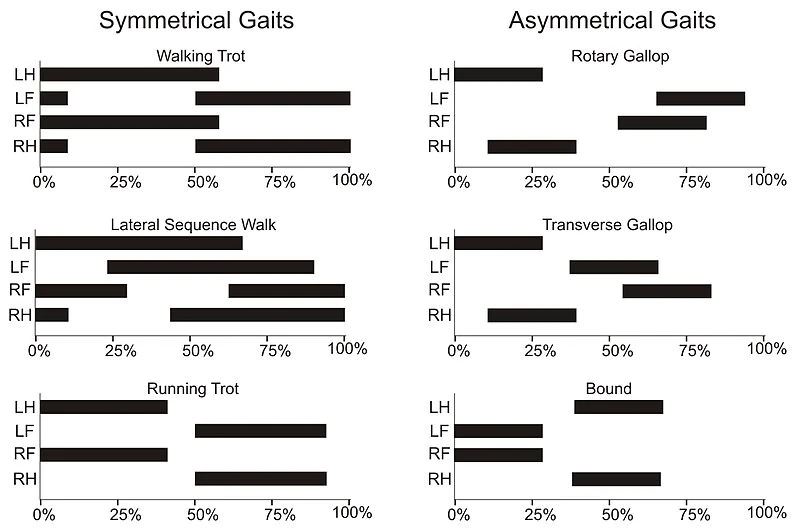

{kind=link}
Graph showing the time each feet touches the ground in different quadrupedal gaits. In symmetrical gaits right and left limbs strike the ground alternately.
Gait is the pattern according to which legs move during locomotion. It can be classified in symmetrical and asymmetrical or leaping (whether movement on one side is followed by one on the opposite side or not), in walking and running (whether at least one foot touches the ground in any moment or not) or according to the number of limbs. Phase is the fraction of a step that passes between the contact of one foot with the ground and the following one (for walking gaits, it's typically (100/N)%, where N is the number of legs). The time of contact of each foot is also expressed as a fraction. (Note: LH = left hind; LF = left front; RF = right front; RH = right hind)
| N | Gait | Phase | Contact | Notes |
|---|---|---|---|---|
| 2 | Walking | 50% | 75% | Both feet touch the ground for half of the time. |
| 2 | Jumping | ~ 0% | 40% | Feet strike and leave the ground at the same time. |
| 4 | Lateral sequence | 25% | 60% | The order of step is: LH, LF, RH, RF. |
| 4 | Walking trot | 50% | 60% | LH and RF move together, and so do LF and RH. |
| 4 | Running trot | 50% | 40% | Similar to the walking trot, but with a small jump. |
| 4 | Amble | 50% | 40% | Legs on the same side move together (e.g. camels). |
| 4 | Gallop | > 50% | 20-30% | Only one foot on the ground for most of the step. |
| 4 | Bound | 50% | 40% | Front feet and hide feet strike the ground together. |
| 6 | Hexapod walk | 30% | 70% | Always 3-4 feet on the ground. |
| 6 | Tripod gait | 50% | 70% | Always three opposite feet on the ground (e.g. insects) |
| 6 | Hexapod run | 20-30% | 40% | 2 feet in the front and 2 in the opposite back on the ground. |
- Brachiation: Some arboreal animals, such as gibbons and orangutans, move between three branches by swinging with their arms (some New World monkeys also use their prehensile tail). The body works as a pendulum (the v ~ √(lg) rule should thus still be valid). Brachiation pattern are simply upside-down gaits. The Furaha Blog names "cernuation" a form of brachiation where the whole body rotates with each swing; this doesn't present mechanical difficulties, but has the problem of eye displacement and rotation.
- Multipedal gaits: Centipedes and millipedes (and Furahan rusps) walk by moving their rows of legs in a wavelike fashion, creating a "cluster" of raised limbs that appears to move forward or backward as the legs are moved in sequence. With a backward ripple, legs clump together on the ground, and remain spaced in air (possibly forcing the animal to move them sideways), while a forward ripple has the opposite effect, so that legs are evenly spaced when they touch the ground.
- Radial gaits: Furaha's spidrid movement page shows two different gaits: having eight legs, one spidrid has a phase of 100/8 = 12.5%, while another moves legs in pairs, and the feet touching the ground always trace a rectangle under it; this post contains different simulations of radial locomotion. Kangaroos have a very peculiar gait in which they move with five appendages, alternating the advancement of the two large feet with that of arms and tail. Finally, the xenohox gazelle runs rotating around its oro-aboral axis, which is parallel to the ground; its three spined-body bears three leg, each of which point skyward before striking the ground.
| Garden snail (Helix aspersa) | 0.10 km/h | 0.84 l/s | Monopod |
| Three-toed sloth (Bradypus tridactylus) | 0.24 km/h | 0.12 l/s | Brach. quadruped |
| Galapagos tortoise (Geochelone nigra) | 0.27 km/h | 0.06 l/s | Quadruped |
| American cockroach (Periplaneta americana) | 2.7 km/h | 19 l/s | Hexapod |
| Water strider (Gerris remigis) | 3.6 km/h | 50 l/s | Water hexapod |
| Gray squirrel (Sciurus carolinensis) | 18 km/h | 18 l/s | Quadruped |
| Human (Homo sapiens) | 24 km/h | 3.7 l/s | Biped |
| Black mamba (Dendroaspis polylepis) | 32 km/h | 2.6 l/s | Nulliped |
| Black iguana (Ctenosaura similis) | 35 km/h | 8.0 l/s | Quadruped |
| African elephant (Loxodonta africana) | 40 km/h | 1.6 l/s | Quadruped |
| Giraffe (Giraffa camelopardalis) | 52 km/h | 2.8 l/s | Ambling quadruped |
| Rabbit (Sylvilagus floridanus) | 56 km/h | 35 l/s | Quadruped |
| Greyhound (Canis familiaris) | 63 km/h | 24 l/s | Quadruped |
| Red kangaroo (Macropus rufus) | 70 km/h | 9.7 l/s | Biped |
| Ostrich (Struthio camelus) | 70 km/h | 7.8 l/s | Biped |
| Horse (Equus caballus) | 88 km/h | 12 l/s | Quadruped |
| Pronghorn (Antilocapra americana) | 98 km/h | 19 l/s | Quadruped |
| Cheetah (Acinonyx jubatus) | 113 km/h | 22 l/s | Quadruped |
Vertical surfaces[]
Liquid surfaces[]
On very small scales, surface tension (the cohesive force of the molecules of a liquid that oppose to external forces) can support the weight of objects denser than the liquid. Among the more likely thalassogens (mercury is not likely to be found in large amount), only molten sulfur provides a surface tension as strong as water, and few others can compete.


{kind=link}
Two water striders (Gerridae) mating. Notice the indentation in water surface.
| Nitrogen | 6.6 mN/m | Cyanogen | 23 mN/m |
| Helium | 9.6 mN/m | Ammonia | 23 mN/m |
| Carbon dioxide | 1-10 mN/m | Sulfur dioxide | 29 mN/m |
| Carbon monoxide | 10 mN/m | Carbon disulfide | 32 mN/m |
| Oxygen | 13 mN/m | Water | 73 mN/m |
| Methane | 14 mN/m | Sulfur | 73 mN/m |
| Hydrogen cyanide | 17 mN/m | Mercury | 470 mN/m |
Since surface tension is measured in units of force per units of length, where the force is parallel to the liquid surface but perpendicular to the length. the upwards force needed to counteract weight is given by threadlike strands, such as the hairs that cover a water strider's feet. A reasonable total length of 100 metres for all the hairs gives 100*(s/1000) newton of force, where s is the surface tension given in the table above; for water and sulfur, 7.3 N; for cyanogen and liquid ammonia, only 2.3 N. Assuming an Earthlike gravity of 9.8 m/s2, this means 7.3/9.8 = 0.744 kg on water and sulfur surface, 2.3/9.8 = 0.235 kg on cyanogen and ammonia.
Since surfactants reduce the surface tension, they can be secreted and used, as the Stenus beetle does, to reduce surface behind, thus creating a net force pushing forward; an aquatic predator could use them to make surface-walking preys fall in the liquid below. What said about land-locomotion limb structure and gaits still applies, keeping in mind that surface-walking works only for very small organisms.
References and readings[]
- Extraterrestrial Biomechanics (Xenology) (also see further pages)
- Mathematical Ideas in Biology (John Maynard Smith, 1968)
- On Growth and Form (D'Arcy Wentworth Thompson, 1917)
- Comparative biomechanics (Steven Vogel, 2003)
- On Being the Right Size (J. B. S. Haldane, 1928)
- A matter of scale
- Biomechanics and The shape of quadrupeds as a function of scale
- Furahan Biology and Allied Matters (many reflections on the mechanics of posture and locomotion)
Notes[]
- ↑ There is at least an exception: the Triassic turtle Odontochelys had only a ventral armour, probably to defend itself when swimming on the surface of the sea.
- ↑ In fact, a balloon full of hydrogen, assuming Earthlike air and a membrane similar to Mylar (0.1 mm thick and 1.2 times as dense as water), is able to lift weigh at all only with a radius just above 30 cm, which means 118 litres of hydrogen. Anything smaller than that and it falls instead of rising, even without a load.