Together with energy gathering and management, reproduction is one of the basic functions of life - in fact, something usually used to define life itself. Evolution is based on the reproductive success of a species, individual or gene: at any level, self-replication is what living systems "want" more than anything else.
Asexual reproduction[]
The most
Binary fission in a bacterium.
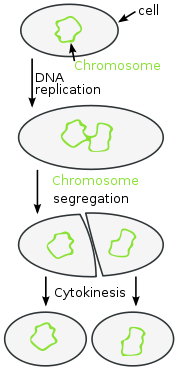
simple form of reproduction (available only to unicellular organisms), and the only one employed by bacteria, is binary fission: the cell simply divides in two parts, each of which receives enough cellular structures and genetic material to survive independently, and grow to the former size. In eukaryotes, before replication the DNA is collected into chromosomes, which are duplicated and split through mitosis.
Gemmation involves the development of new individuals from undifferentiated tissue on the parent's body. Besides unicellular organisms, it's also known in animals, but only very simple ones, such as sponges and hydrae (see image). Sponges also can reproduce through gemmules, cellular mats that rebuild the whole body in a new location.
Similar to gemmation, fragmentation requires a small part of an organism to be separated from the rest of the body, and regrow it until it becomes a complete individual again. This method, of course, also requires a very simple body organization. All fungi and moulds can grow from a small fragment of mycelium, while many plants, such as moss, cacti and willows can grow respectively from thallus, parts of stem and branches. Among animals, annelids, flatworms, sea anemones, corals and starfish can reproduce in this way: in architomy, the separation occurs along a symmetry axis, and regeneration is specular; in paratomy, the two halves are different, and the regeneration is preceded by the pre-generation of the missing parts.

Gemmation in Hydra oligactis, a freshwater cnidarian.
Vegetative reproduction (or clonation in a strict sense) is common in plants, even those otherwise able to reproduce sexually: similar to gemmation, meristematic cells can differentiate and produce new structures, allowing the plant to expand quickly, feeding the clones until their complete self-sufficiency, often through a lateral stem (stolon). Other plants can regrow from buried tubers or bulbs, that also work as food storages.
Agamogenesis is any form of reproduction that involves gametes but not fecundation. The development of an unfertilized egg into an embryo and then an adult individual is specifically called parthenogenesis; since an egg is the only gamete that contains enough food and structures for later development, the parthenogenetic sex is always a female. In apomictic parthenogenesis eggs are produced by mitosis, and the offspring is always a clone of the mother; that's the case for aphids and many flowering plants. In automictic parthenogenesis, eggs undergo meiosis (see below), in which chromosomes are duplicated and recombined, allowing a limited genetic variation between the mother and the offspring. Parthenogenesis is connected to polyembryony (multiplication of an embryo in several identical twins, common in armadillos and wasps) and pseudogamy (where the presence of pollen or spermatozoa triggers the embryonal development but does not contribute genetically). Among vertebrates, parthenogenesis is known in many species of reptiles and sharks. Some of them (a blind snake, a gecko and a lizard) are always parthenogenetic and lack males.
Sexual reproduction[]
Asexual reproduction is, generally, faster and more efficient than sexual reproduction: even a lone individual is able to
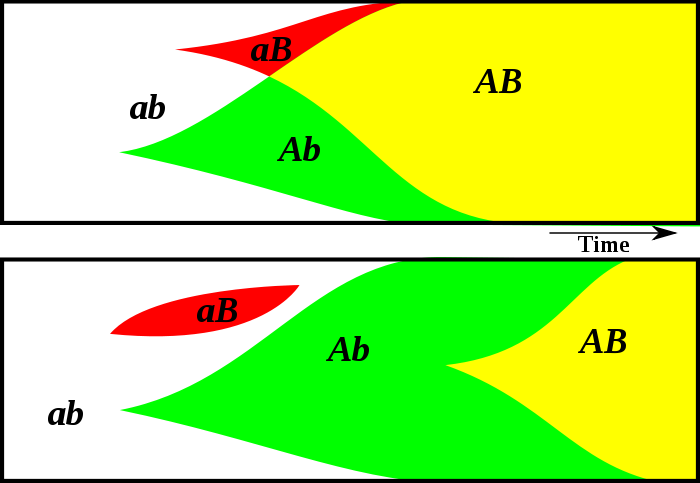
Sex (above) creates new genotypes faster than asexual mutations (below).
produce offspring, rather than requiring the presence of others, and offers at it the entire parenthood of the offspring, rather than having to share it with others (and thus having to spend energy to raise descendants that also carry unrelated genes). This problem is amplified in organisms where only one parent is capable of bearing young.
However, fast, asexual reproduction can produce only clones of the parent, barring occasional mutations; sex greatly accelerates evolution recombining genotypes and thus spreading genes from one lineage to another. In fact, while hydrae and aphids reproduce asexually most of the year, they switch to sexual reproduction in early winter, when a greater genetic variation is needed to survive the harsh weather.
According to the Red Queen Hypothesis, sex provides an advantage in fighting parasites, that usually have very short life cycles and thus evolve quickly; if the parasites present different forms, they're likely to change cyclically, and represent to their hosts the same challenges in different times. For this reason, the way sexual reproduction preserves different genotypes allows them to resurface every time they're advantaged against particular parasites.
Sexes and gametes[]
In sexual pluricellular organisms the transport of genes is entrusted to highly specialized cells, called gametes (that usually carry only part of the individual's genes; see below). A number of gametes (almost always two) join together in fecundation, forming a zygote which then develops into a fully-grown individual. Sexual reproduction requires a certain distinction of genotypes (sexes or genders). In isogamous species, this distinction has no morphological consequences, and neither sex can be called "male" or "female", but they're rather identified as "+" and "-" if both gametes are mobile (as in Chlamydomonas) and as "a" and "α" if neither are (as in yeasts).
When two sexes are clearly distinct by the nature of their gametes, individuals with small, numerous and mobile gametes are called males, and those with few large and static gametes are called females. This distinction probably derives from a double specialization where female gametes have been selected for better support structures and nutrient storage; male gametes didn't thus need to carry nutrients with them and were free to specialize in speed, often adding movement structure such as the flagellum of spermatozoa.
Genetics of sex[]
Chromosomes[]
DNA is stored inside cells wrapped around proteins (histones), forming structures called chromosomes. During mitosis, each chromosome duplicates, becoming X-shaped; the two identical halves (chromatidia) are then divided between the two cells that result from the mitosis process. Since each gamete carries half of the genome of the individual, sexual species have a double genome formed by a number of chromosome pairs; such species are called diploid, as each somatic cell (part of the body) contains two copies of each chromosome, while their gametes contain only one, and are thus haploid. This means that each gene, in a diploid organism, can have two different interacting varieties,
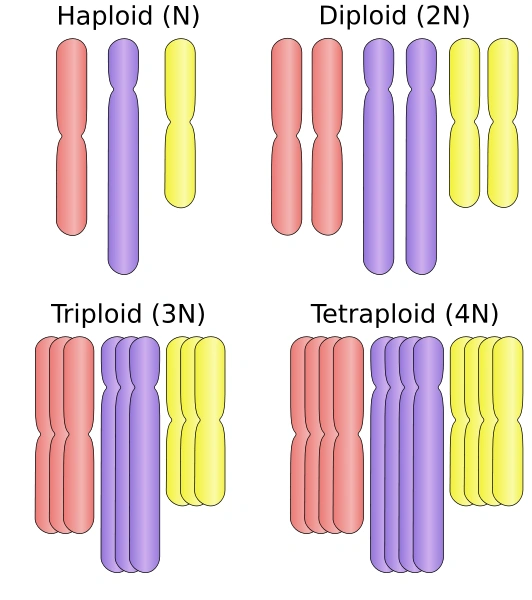
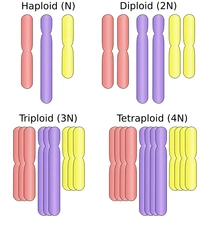
The most common ploidies.
called alleles.
More generally, the number of copies is called ploidy. The number of chromosome pairs/groups can vary from one (in jack jumper ants) to 631 (adder's-tongue ferns), while humans have 23 (and therefore, being diploid, 46 chromosomes). While gametes, barring malformations, are always haploid, the cells can develop by duplication a higher ploidy: some tardigrades and watermelons are triploid (3 copies per chromosome), trouts and salmons are tetraploid (4 copies), kenai birches are pentaploid (5), kiwis and wheat are hexaploid (6), dahlias and sturgeons octoploid (8), some strawberries decaploid (10); the plant Celosia argentea and the frog Xenopus ruwenzoriensis reach dodecaploidy (12 copies of each chromosome). However, the vast majority of eukaryotes are diploid.
| Species | Chromosomes | Species | Chromosomes |
|---|---|---|---|
| (all prokaryotes) | 1 | Wheat, rat | 42 |
| Jack jumper ant | 2 | Rabbit, hamster | 44 |
| Anopheles mosquito | 6 | Human, guppy | 46 |
| Fruit fly | 8 | Gorilla, chimpanzee | 48 |
| Arabidopsis thaliana | 10 | Elephant, silkworm | 56 |
| Barley, rye, pea | 14 | Horse, chinchilla | 64 |
| Pigeon, cabbage | 18 | Dog, wolf, coyote, chicken | 78 |
| Maize | 20 | Carp, goldfish | 104 |
| Beer yeast | 32 | Kingfisher | 132 |
| Cat, otter, pig | 38 | Adder's-tongue fern | 1262 |
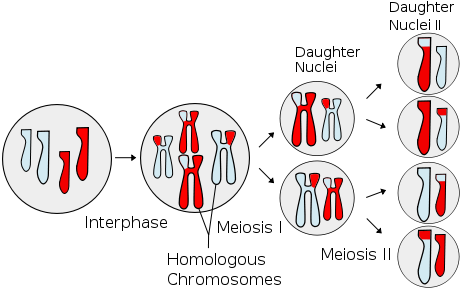
Scheme of meiosis. In this image, different chromosomes are identified by size, while the colour reveals the parent they come from.
The production of haploid gametes requires a process called meiosis: it's similar to mitosis, but it contains an exchange of genes between homologous chromosomes (that is, belonging to the same pair/group); it divided in meiosis I, where chromosome pairs are divided between two diploid daughter cells, and meiosis II, where the chromosomes themselves are split in their chromatidia (see here an animation of the process). The end result is four haploid cells, each of them containing one chromatidium for each original chromosome pair. Each of them contains genes from both parents, thanks to the previous recombination: all the final cells are genetically unique.
Life cycle[]
The cyclic change of ploidy among the generations of a species are summarized in the biological life cycle:
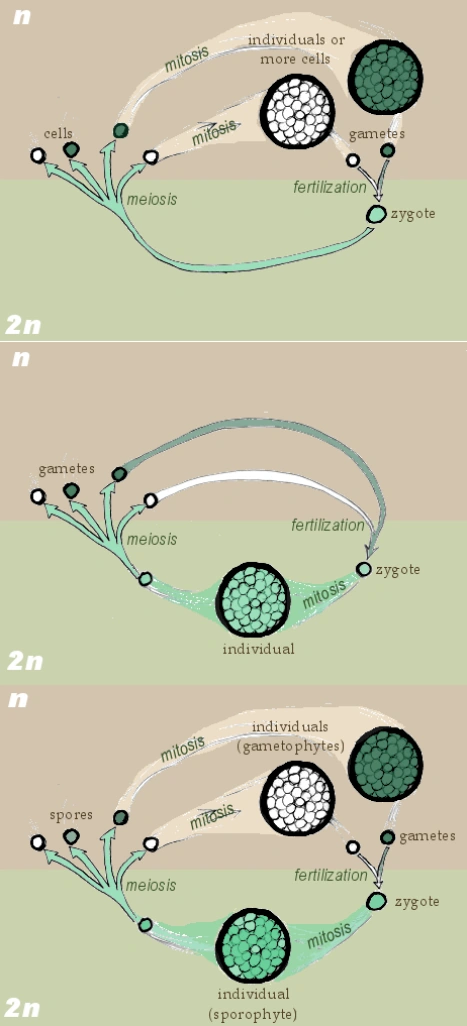
Three forms of biological life cycles (succession of all the forms or generation of a particular organism). From top to bottom: haplontic life cycle, with zygotic meiosis; diplontic life cycle, with gametic meiosis; haplodiplontic life cycle, with sporic meiosis.
- Haplontic life cycle: the fertilized zygote (diploid) undergoes immediately zygotic meiosis, producing a number of haploid embryos that still have the genetic traits of both parents. The adult individuals that grow from them are still haploid and produce gametes by simple mitosis: male ones in the antheridium and female ones in the archegonium. This life cycle is found in many algae, protozoans and most fungi.
- Diplontic life cycle: the zygote multiplies by mitosis, forming the diploid body of an adult organism. This produces by gametic meiosis haploid gametes: male ones in stamina or testicles and female ones in the ovaries. All terran animals follow this scheme, as well as brown algae and yeasts.
- Haplodiplontic life cycle (also called diplohaplontic, diplobiontic or dibiontic): there are alternate generations of haploid and diploid organisms. As in the diplontic cycle, the zygote multiplies by mitosis in a diploid individual, the sporophyte. This creates, through sporic meiosis, haploid spores that, without fecundation, multiply by mitosis into a new, haploid, individual, the gametophyte, which produces haploid gametes via mitosis. This cycle exists in most plants and some fungi. Usually, one of the generation prevails on the other: mosses have a dominant haploid generation, so that the sporophyte appears to be a mere organ of the gametophyte, despite its genetic diversity, while in ferns the sporophytes are the only recognisable individuals. In seed plants, the entire pluricellular body is a diploid sporophyte, except for pollen and the contents of the ovule, which are very small gametophytes. "Animal" life on Ilion follows a similar scheme: moving individuals are haploid, while their "uterus" is diploid.
Sex determination[]
Often, the sex of an individual depends from a particular chromosome pair, that exists in two varieties: one sex (homogametic) has two copies of the same type of these, while the other (heterogametic) has different chromosomes, and thus its gametes can carry two different sexual chromosomes rather than always the same. In mammals, some insects (such as fruit flies) and plants (such as ginkgos), females are homogametic (XX chromosomes) and males heterogametic (XY chromosomes). This is the XY system. The opposite occurs in the ZW system, where males are homogametic (ZZ) and females heterogametic (ZW), as in birds, many fish and crustaceans, insects such as butterflies and some reptiles (such as monitor lizards).
Many insects use the X0 system, where females have two sexual chromosomes (XX) and males have only one (X). This happens in eusocial insects such as ants and bees, where queens produce females from fertilized, diploid, eggs and males from unfertilized, haploid, eggs (if female jack jumper ants have two chromosomes, males have only one at all). The southern platyfish has an even more complex system, with three sexual alleles (W, X, Y) that allows WY, WX or XX as females, and XY or YY as males. The platypus has five pairs of sexual chromosomes, the workings of which are still not entirely understood.
There are examples of environmental sex determination: in alligators, tuataras, turtles and some birds it depends from the temperature of incubation of eggs, that influences the embryo's hormone production. In other cases, the sex can be determined by other hormonal influences or even bacterial infections, such as Wolbachia for many arthropods; several organisms functionally change their sex during their life (see below).
Variations on sexes[]
While the most common form of sexual reproduction involves two diploid parents of different sexes, life on Earth has experimented with many variations.
Number of sexes[]
In hermaphroditic species, each individual produces both types of gametes, so, while sexual reproduction still occurs, there's functionally only one sex. It's useful for species, such as slugs, with low population density and/or unable to travel for a great distance, as it ensures that any partner a particular individual meets will be apt for procreation. Hermaphrodites also include planarians, earthworms, sponges, hydrae and many fish; they can have gamete-producing glands of both types or only one (ovotestis), that produces both eggs and spermatozoa.
The presence of both types of gametes in the same individuals allows autogamy, or self-fecundation, in which the same individual has the role of both parents. The process of meiosis and following recombination allows a certain genetic variety in the offspring anyway. While most animals have only one reproductive system, land plants have multiple sexual organs (flower), which usually contain both male and female structures, making autogamy the rule in the vegetable kingdom; however, about 14% of plant species have only structures of one sex in each flowers, and they're either monoecious (both kinds of flower on the same plant) or dioecious (only one kind of flower on each plant); see full terminology here.
In the opposite case, the number of sexes can increase beyond two (multisexuality): it can happen, for example, that an organism has three sexes, and any combination of two can procreate. In this case, two thirds of the individual met can be sexual partners rather than only half; this happens in the protozoon Tetrahymena thermophila, which has seven sexes, each of which can mate with any else but its own. The slime mold Physarum polycephalum has three genes that control its sex: MATa has thirteen possible alleles, MATb also thirteen and MATc three, which allows for 507 different sexes, each of which can mate with a specific set of others.
In all these cases two sexes among the possible ones are needed for reproduction, but a species can be imagined where three sexes together are required, fusing three forms of gametes. This, however, actually reduces the chance that all the compatible organisms meet, and that they're all free from genetic defects.
Permanence of sex[]
Even if two sexes are present, they don't necessarily remain fixed on the individuals: a species can ensure the benefits of hermaphroditism while keeping specialized sexes through sequential hermaphroditism, common among fish, mollusks and plants (for which it's called dichogamy), where an individual naturally changes its sex at a certain point of its life. In the amphipod crustacean Gammarus pulex, all individuals, born sexless, grow as males; once they exhaust the spermatozoa, their ovaries fully mature, and they become fertile females. In Xiphophorus fish, the opposite occurs: each individual becomes a male after having exhausted its eggs. The male-to-female transformation is called protandry, the female-to-male one protogyny; the last one is far more common, as eggs requires more resources than spermatozoa and thus they're more restricted in number.
Heterogony is a combination of sexual reproduction and parthenogenesis, found in aphids, rotifers and water fleas: a population expands quickly through parthenogenesis, but when adverse conditions occur, such as winter or a food shortage, females produce by automictic parthenogenesis a number of males, which then they mate with; sexual reproduction helps thus to increase genetic variety. The sporophyte/gametophyte cycle is a similar process, with other variations: red algae have two diploid generations, the carposporophyte and the tetrasporophyte. Apicomplexa protists (such as the malaria plasmodium) have yet more complex life cycles.
Eusociality[]
Also see Introduction to Eusociality, Genetic sentience, Social organization
In eusociality, all the adults of a particular generation live together and raise the offspring produced by a small group of fertile individuals. Eusociality is found in several groups of insects, such as hymenopters (ants, some bees and wasps), termites, some Hemiptera, the australian weevil Austroplatypus incompertus, the shrimp Synalpheus regalis and, among vertebrates, only two species of mole rats. In the most advanced forms of eusociality, as in ants and termites, there is a morphological specialization of the different castes, including sterile ones (workers, soldiers). This means that most individuals, rather than raising their own offspring, help raise their mother's or sister's. Often, eusocial species live in a sort of "fortress" collectively defended; honeybees, termites and paper wasps build their own nest (respectively with wax, dirt and chewed plant matter), while Synalpheus inhabites sponge reefs.
With a two-sex diploid system, each son receives half of its genome from each parent, and thus has a ½ kinship with both; it shares half of the father's genes and half of the mother's genes with a sibling, with which it has still a ½ kinship. Ants and bees have diploid females and haploid males (as they're born from un-fertilized eggs): that means that the parent/son kinship is still ½, the sibling/sibling kinship is ¾ (two sister ants share half of the mother's genes and all of the father's genes). Because of this, an ant can better spread its genes by raising its siblings, the queen's offspring, rather than its own.
Even in all-diploid organisms, though, a strictly monogamous queen has sons that are as related with each other as they are with their own offspring, so that in a situation that requires, for example, a limited growth, raising the queen's offspring can, again, be convenient. This is believed to be the case for termites and Synalpheus. Besides, eusociality's low dispersion of genes is favoured in a species that depends from sparse resources that require collective defense, such as the sponges' water intake for Synalpheus and underground tubers for mole rats (see here). Eusocial vertebrates are a staple of speculative biology: see TFIW's spink, The New Dinosaurs's gestalt and Man after Man's hivers.
Physiology of reproduction[]
Fecundation[]
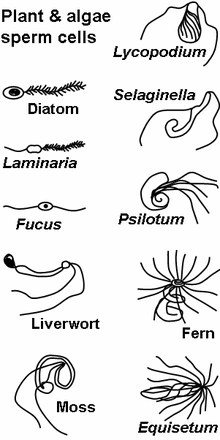
Spermatozoa of various plant and algae species.
Fecundation or syngamy is the process of fusion of two haploid gametes to produce a diploid zygote (whose ploidy can increase further after that by chromosome duplication). A fecundation where more gametes are involved (say, three haploid gametes into a triploid zygote) could be imagined for different planets, though they make mating more difficult and unlikely.
In aquatic lifeforms (or anyway living in a dense medium), eggs and spermatozoa can simply be released in water, though this requires a great volume of gametes for each successful fecundation; in many cases (as in octopi, amphibians and most fish) the male fertilizes the eggs just after they've been laid to ensure the fecundation. Out of water, the organisms need to ensure that gametes are protected from dehydration and brought to contact; various terrestrial animals enclose spermatozoa in a waterproof proteic capsule (spermatophore), while flower plants disperse capsules that produce them directly inside the female reproductive organs (pollen).
Internal fecundation[]


Shark pterygopods, that form a channel to direct sperm.
On dry land, it's common internal fecundation, that is, the fecundation of eggs still inside the female body. This allows a better control of the offspring's parenthood and cancels the necessity of preserving and transporting gametes outside the body. The males of several animal groups have developed copulatory organs for this purpose: rotifers have ducts made up by hair cells; more complex structures have been derived from limbs (that myriapods use to manipulate the spermatophore), from pedipalps (that spiders use to deposit them on a silk strand), from pelvic fins (the gonopodium of several bony fish and the pterygopods of sharks), from tentacles (the hectocotylus of cephalopods), from extensions of the abdomen (the edeagus of insects) or of the cloaca (the penis of mammals, reptiles and tailed frogs). In amphibians and most birds, the transfer of gametes simply occurs by contact among the two cloacas (cloacal kiss).
Many slugs have "love darts" (or gypsobela), calcareous or chitinous spikes that they can shoot into another slug's body; while they're not used to transfer sperm, such a system might be employed by alien lifeforms. In argonauts, a kind of swimming octopuses, the hectocotylus detaches and swims on its own towards the female to fecundate it.
Pollination[]
Since internal fecundation requires the mobility of organisms (though there are sessile barnacles that mate with a penis several times longer than the rest of the body), land plants transport their pollen exploiting several different environmental factors: the complex of adaptations developed for this purpose is called pollination syndrome.
- Roughly the 80% of plants has its pollen carried by animals that bring it directly to another flower (biotic pollination), reducing dispersion and waste. Usually, the pollinator receives food from the flowers (nectar).
- Entomophily: the plant entrusts its pollen to insects such as bees (melittophily), butterflies (psychophily), moths (phalaenophily), flies (myophily) and beetles (cantharophily). Melittophile flowers are often yellow, white or blue, with elaborate patterns visible only with ultraviolet; psychophile ones are large and pale, with "landing pads" for the butterflies, with pollen and nectar hidden inside long and narrow tubes; sapromyophiles ones (which rely on blowflies, such as Rafflesia) are very large, hollow, reddish and smell like feces or rotting meat; cantharophile ones are wide and flat, with easily accessible pollen, and smell like fruit and rotting leaves.
- Ornithophily: pollinating birds include hummingbirds, sunbirds and honeycreepers. Ornithophile flowers are usually red and rich in nectar, and often have a support for the bird to stand on.
- Zoophily: pollinating mammals include monkeys, lemurs, possums, bears, deer and rodents, but the most important ones are by far fruit bats (chiropterophily); chiropterophile flowers are large, white or otherwise light-coloured and bell-shaped, with large pollen grains; they often open by night. There are also examples of pollinating reptiles, such as the gecko Hoplodactylus from New Zealand.
- Of the remaining plants that rely on abiotic pollination, the 98% uses wind (anemophily). Anemophile flowers produce an enormous amount of pollen, often in very small grains and with long and ropy stigmata that catch other flowers' pollen from the air. Conifers and many grasses are usually anemophile.
- The rarest form of pollination is hydrophily, which relies on water. The adaptations are very similar to those for anemophily; it's found in aquatic flowering plants such as eelgrass or posidonias.
Dissemination[]
In the case of sessile species with internal fecundation or self-fertilizing, like many trees, the propagation of genes can be ensured through the dispersion of zygotes (seeds in the case of conifers and flowering plants). In many cases, the fruit (called dehiscent) open spontaneously once ripe to release its seeds.


The anemochore, dehiscent crepe myrtle (Lagerstroemia sp.) opens to release its seeds.
- Barochory: the seeds simply fall by gravity from the plant. This method works best, of course, for trees (such as apple trees and coconut palms) or epiphytes (such as the passion flowers). Bean pods are dehiscent fruits that open to let the seeds fall out.
- Boleoautochory: the seed is forcefully expelled from the fruit, as in the exploding cucumber, that squirts out a mucilaginous fluid that contains the seeds (see a video here). The Pilobolus mould, that grows on herbivore dung, high less than 1 cm, can throw spores up to 2 m away.
- Herpautochory: the seeds slide out from the open fruit thanks to trichomes (hair-like surface growths) that allow movement in only one direction. Many ears of grain have a similar feature.
- Anemochory: the smallest and lightest seeds are transported by wind. Dandelion seeds have a feathery structure (pappus) that works as a parachute; maples and elms produce samaras, small winged fruit that glide for long distances. This method is common in open habitats such as plains and dry forests.
- Hydrochory: mangrove fruits germinate if they fall on the ground during the low tide, otherwise they're drawn back in the sea and carried by the current; in water lilies, the seed sinks slowly to germinate underwater; coconuts can travel for thousands of kilometers carried by sea currents.
- Zoochory: the seeds are carried by animals (or other mobile organisms).


Evident elaiosomes on Afzelia africana seeds.
- Epizoochory: the seeds adhere to the animal's surface with spiny or hooked trichomes. This method, albeit generally rare, is commonly found in the carrot and sunflower families. Of course, it works best if the animals have a hairy or feathery cover rather than a smooth skin.
- Endozoochory: the fruit are ingested by an animal and the seeds are expelled with the feces. In tropical rainforests, endozoochore plants form up to the 95% of the flora; they have sugar-rich fruit, often with bright colours such as red and yellow. The most important animals for endozoochory include parrots, toucans, hornbills, tapirs, orangutans and chimpanzees.
- Squirrels and jays hide and store acorns, many of which germinate if they're not found; this allows oaks to propagate their genes. Myrmecochory is a variety, relying on ants, that evolved perhaps a hundred times, in the 9% of modern plant species. Such plants (myrtle, spurges, hyacinth) develop elaiosomes, oil-rich structures: to eat these, ants carry the seeds in the nest, where they can germinate.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
One can imagine an even wider variety of dissemination methods; mobile fruits that crawl on the ground with functional muscles (a similar situation happens on Ilion), or that float in a dense atmosphere with lighter-than-air balloons (with the restrictions detailed here).
Serotiny is an adaptation in which the seed release is triggered by an external event, such as the death of the parent plant (necriscence), moisture (hygriscence), warming by the Sun (soliscence), drought (xeriscence) or even fire (pyriscence). Eucalyptus trees are pyriscent: they produce a highly flammable sap that purposefully feeds wildfires to destroy other plants and create space for the seeds.
Gestation[]
After fertilization, the egg has to develop until the embryo is ready to hatch. Completely formed eggs develop a structure (yolk) containing a large amount of proteins and lipids to nourish the growing embryo (which makes eggs a very attractive food for many predators).
- Ovuliparity: the females lays unfertilized eggs, and fecundation occurs after that. This happens in arthropods, fish and most amphibians. Zygotes develop on the outside; in fish and amphibians, the young carry with them the yolk until it's completely depleted. This is the only form of gestation that doens't require internal fertilization.


{kind=link}
Eggs of various birds, reptiles, fish and invertebrates.
- Oviparity: after internal fecundation, the zygotes develop inside the female's body into fertilized eggs. This strategy, most common among reptiles and birds, allows the egg to grow an external shell to protect itself from predators or dehydration, since male gametes don't need to penetrate a mature egg.
- Ovoviviparity: the fertilized egg is not laid but remains inside the female body (that doesn't interact with it in other ways) until it hatches. In this case, the egg usually doesn't have a shell. This happens in many lizards and snakes; the hermaphrodite leech Marsupiobdella africana and male sea horses keep their eggs in a ventral pouch; in the gastric-brooding frog and the Darwin's frog, eggs hatch respectively in the stomach and in the mouth. Several insects, such as tsetse flies give birth to alive larvae (ovolarviparity).
- Histotrophic viviparity: zygotes develop inside the female oviduct, where embryos are fed by oophagy (eating unfertilized eggs), adelphophagy (eating other embryos, as in many sharks and in the alpine salamandra), mucoid histotrophy (eating yolk or mucus, as in dogfish and electric rays) or lipid histotrophy (eating specialized lipid-rich structures or trophonemata, as in stingrays).
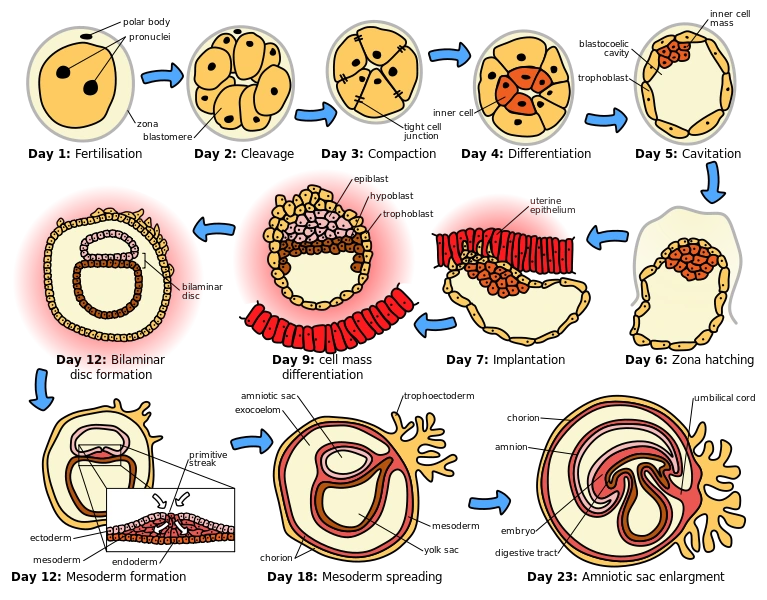
{kind=link}
Scheme of the human embryonic development in the first month.
- Hemotrophic viviparity: zygotes develop inside the female oviduct, and embryos are fed by a direct link with the mother's bloodstream. This is typical of placental mammals, where the link occurs through the placenta, while the frog Gastrotheca ovifera uses specialized gills. The grass Poa alpina can also be considered viviparous, as seeds detach from the parent's body after germination, as the unnamed future plants found with Dixon's unknown creature.
| Species | Days | Species | Days |
|---|---|---|---|
| Opossum | 12-13 | Lion | 108 |
| Pigeon | 11-19 | Pig | 115 |
| Budgerigar | 17-20 | Sheep | 145 |
| Mouse | 18-20 | Baboon | 187 |
| Chicken | 22 | Human | 259-264 |
| Rabbit | 33 | Horse | 336 |
| Kangaroo | 32-39 | Giraffe | 420-450 |
| Fox | 51-63 | Rhinoceros | 487 |
| Dog | 62 | Sperm whale | 480-590 |
| Cat | 65 | Elephant | 600-660 |
Parental care[]
Several species of birds, arachnids, all mammals and even the leech Helobdella striata feed their young as part of their reproductive strategy. Food is generally the same that the parent usually eat, often previously chewed or digestes (wolves, vampire bats and most birds resort to trophallaxis, regurgitating the food in the young's mouth). Among fish, amphibians and insects are common the trophic eggs, unfertilized eggs that are laid so that the hatched offspring can feed on them.
In some cases, the parent(s) can produce a nourishing substance: the most obvious example is the milk of mammals, a liquid rich in calcium, lipids and proteins, produced by the females' mammary glands. In monotremes, milk is secreted as sweat through skin ducts, while in other mammals the ducts' openings are concentrated in the nipples. The bat Dyacopterus spadiceus is the only known mammal where milk is also produced by males.
Some birds (flamingos, pigeons and penguins) also produce a nourishing, semisolid paste (crop milk) secreted in the throat of both parents (only males in pigeons) and regurgitated in the offspring's mouth; it allows to survive the lack of animal preys, especially during the development of the flamingo's filterer beak (see here). Discus cichlids of both sexes produce from their skin a mucus their larvae feed on.
Since the young develop in the female's body, parental care is usually provided by the female as well, especially in mammals; most exceptions are found in birds, where the long-lasting eggs allows a more varied behaviour. Eggs are incubated only by females in hummingbirds, birds of prey and passerines (often with assistance from the male); more strictly monogamous birds, such as cormorants, finches and woodpeckers, share the incubation; in birds where the female mates with several males, such as jacanas and phalaropes, the eggs are incubated by the male. In most cases, invertebrates don't look after their offspring at all, with some exceptions - such as the sand wasp Bembix rostrata, that feeds a few larvae with freshly killed preys.
References[]
- Alien Sex (Xenology) (also see further pages)