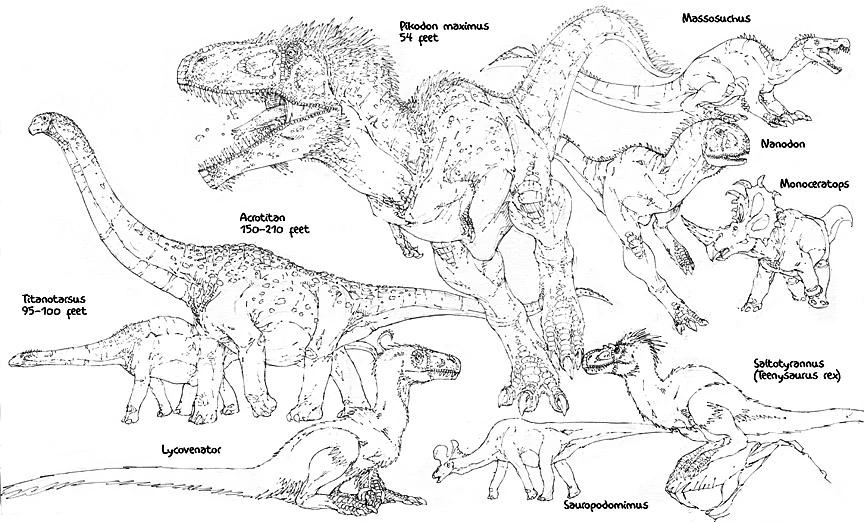
Examples of Extinct Species in the Spec World:
- Sauropodimimus giganticus (Late Oligocene) (Chattian) Asian lambeosaurid that attained enormous sizes, growing up to lengths of 16 meters. One of the last of the lambeosaurs with the extinction of Sauropodimimus and it's kin, Asia was devoid of hadrosaurs until repopulated by stock from North America during the Late Miocene and Early Pliocene. However, recent expeditions back to Spec support the existance of lambeosaur-like creatures living in the Southern Hemisphere.
- Saltotyrannus nanus: (Early Eocene) Also known as "Teenysaurus rex", this tiny, meter long tyrannosaur, also from the Messel Shales is considered by many to be the first true errosaurid (or at least a "proerrosaurid"). The superlative preservation of the Messel shows that this animal possessed the full body-covering of feathers present in its modern cousins.
- Lycovenator messelensis (Early Eocene): A four meter long plesiomorphic dromaeosaur from the Messel Shales putative ancestor for the entire Tertiary Holarctic drak radiation.
- Monoceratops acutus (Late Plestociene): Growing up to lengths of six meters long, Monoceratops was one of many spectacular large North American dinoceratopsians that died out in the Late Pleistocene, due to climate change and ironically, the return of a few possible species of ceratopsians to North America. This issue is still under debate. Monoceratops is a perfect case of convergent evolution due to its appearance of a long extinct centrosaur.
- Nanodon marshalensis (Pliocene): One of the last of the South American abelisaurids from the Pliocene, which grew up to lengths of seven meters long. This creature became extinct shortly after the invasion of northern predators, most notably dromaeosaurs and tyrannosaurs, across the Central American isthmus.
- Acrotitan colossicus: (Oligocene) (Chattian) Neotropical saltasaurid. Amongst the largest and the last of the South American titanosaurs, growing over a mind-boggling 150-200 feet long.The only sauropod left in that area is the Centeotl.
- Titanotarsus steveirwini: (Eocene) (Lutetian-Priabonian) Australian/Antarctic titanosaur. Amongst the last sauropods found in Australia and Antarctica. The reason it went extinct was due to climate change and competition from other herbivores. The only species of Titanosaur left in the area are the Navigatosaurus sp.
- Pikodon maximus: (Miocene) One of the last of the Australian torvodonts and one of the largest terrestrial predators even discovered, at 50 feet in length.
- Massosuchus europensis: (Late Eocene) A European spinosaur, which grew up to lengths of 30 feet long, with a perculiar arm/leg lenghth ratio that suggests a quadripedal stance.
A QUICK INTRODUCTION TO SPECPALEONTOLOGY[]
The history of Spec as a timeline distinct from that of Arel begins roughly 65 million years ago. On Arel, this point in time corresponds with the end of the Mesozoic Era and the dawn of the Cenozoic - the so called Cretaceous-Tertiary (K-T) boundary, although many workers now prefer to divide the Tertiary into the Paleogene(Paleocene-Oligocene) and Neogene(everything else).
Specworld fossils dating from before this boundary most often belong to organisms described in our native timeline’s fossil record, indeed some famous pre-Cenozoic Arel fossil sites such as the Dinosaur National Monument and the Burgess Shales have identical Specworld counterparts that have produced matching specimens. A few Mesozoic Spec fossils (most notably Mirabilotheridium , a monotreme from Cretaceous South America and Lepelara, an early angiosperm) represent new taxa. All these novel finds fit comfortably within the accepted theories of our home-Earth’s Mesozoic history. In all likelihood, these taxa simply represent fossils of creatures that existed on Arel but have yet to be found.
Above K-T boundary however, Spec's fossil record diverges radically from the natural history of our home timeline. The most obvious feature is the fact that the dinosaurs did not go extinct along with a host of other organisms that vanished at the K-T boundary on Arel. Secondly, a great variety of animals that appeared in the early Cenozoic of Arel (particularly large mammals) are missing - without a big K-T extinction event to clear out the ecological playing-field these forms never had the opportunity to evolve on Spec.
"K-T" STRATIGRAPHY[]
Most Spec-researchers continue to use a stratigraphic system based on the one developed and calibrated on Arel. This is feasible as the broad geological and climatic histories of our two worlds, with the exception of the K-T boundary, appear to be identical. For example, the timing of the appearance and disappearance of the index fossils (various marine planktonic forms) that help to demarcate the different stages of the Cenozoic show a strong correspondance between the two timelines. (Note: the actual makeup of the Cenozoic planktonic floras of Spec and Arel are very different, however the arrival and extinction of stratigraphically useful taxa happens at about the same time).
However, there remains one glaring geological difference that separates Spec from Arel - 65 million year old deposits on Spec show no sign of a large impact event or the subsequent extinctions caused by it. In fact, discounting the long-term decline of certain lineages and a drop in marine tropical diversity, there appear to be no abrupt changes in the global biota between about 71 million to 63 million years ago. This roughly corresponds on Arel stratigraphic charts with the start of the Maastrichtian Age (the final stage of the Late Cretaceous Epoch) and the end of the Danian Age (the first stage of the Paleocene Epoch).
Especially noticeable is the continued formation of chalk deposits during this time whereas on Arel, all production suddenly ceases due to the extinction of radiolarians (chalk-producing planktonic organisms). Just as striking is the complete absense of the famous band of iridium-rich deposits, the ominous global signature of the K-T impact.
As a result, some researchers have proposed a number of amendments to stratigraphic table of Specworld based on striking non-conformities in the Specworld fossil record when compared with that of Arel.
- The end of the Cretaceous Period (and thus the Mesozoic Era as well) on Spec is pulled forwards from c.65 million years ago to c.55 million years ago, corresponding to the end of the Paleocene Epoch on Arel. The retention of a distinct Paleocene unit as the first part of the Cenozoic should be abolished.
- The period of time on Specworld corresponding to the Danian Age of the Paleocene on Arel is now incorporated into the Maastrichtian Age of the Cretaceous. Thus the Maastrichtian, which extends from 72-65 million years ago on Arel, extends from 72-62 million years ago on Spec.
- The other subdivision of the Arel’s Paleocene, the Selandian (extending from 62-55 million years ago) is retained as a distinct unit owing to changes in planktonic assemblages. It is however, considered to be the final age of the Cretaceous Period rather than as part of the Paleogene Period.
FROM 65,000,000 YEARS AGO TO THE PRESENT DAY[]
The following account is based on our current understanding of the history of Specworld from the what would have been the end of the Cretaceous on Arel up to the present day. The reader should note that many facets of Spec's fascinating past are still unravelling and that some details are still poorly understood and the subject of intense scientific debate (as well as some rather crude discourses in certain internet forums).
LATEST MAASTRICHTIAN - EARLY PALEOCENE (65,000,000 - 62,000,000 b.p.)[]

Across the globe, the Earth has become a grand stage for spectacular tectonic activity. Great mountain ranges are being pushed upwards accompanied by volcanic activity the likes of which have never been seen before in the Mesozoic. The grandest spectacle takes place on the Gondwanan fragment of India as it dashes itself against the Asian plate. Erupting in their full glory are the volcanoes that will create the Deccan Traps, leaving an area of 1.5 million square kilometers buried under 2 kilometers of flow basalt.
This is not the only upheaval facing the world. Across the globe the oceans have been in retreat for the past few million years, exposing vast tracts of shallow seafloor. Great Britain rises from its watery prison. The two halves of North America are reunited as the Interior Seaway vanishes. The extent of the continental landmasses is dramatically enlarged. Soon it is possible to walk around the entire Northern Hemisphere without getting one's feet wet.
All this activity has put life on Earth under intense stress. Great clouds of volcanic ash screen the sunlight with dire consequences for many phototrophic organisms. Whole continental shelf communities are left high and dry. Once lush island paradises are suddenly faced with continental climates that bring extremes of temperature and erratic rainfall, now often laced with sulphuric acid.
In the oceans, global oceanic cooling and the draining of inland seas leads to a near total annihilation of the world's tropical reef ecosystems. Many once diverse families are eliminated or reduced to a few cosmopolitan species. Entire food chains collapse, from tiny plankton to the apex predators.
The mosasaurs and ammonoids somehow manage to pull through. The vast majority of the graceful elasmosaurs and pliosaurs are not so fortunate, though the elasmosaurs had more luck than the pliosaurs. On the seafloor, once vast beds of giant inoceramid clams dwindle then vanish whilst their cousins, the rudists, are reduced to a handful of coolwater species. Although the rudists as a group will survive up until the present day, their time as the dominant reef-builders is over.
Life on land is also under siege. Death in different parts of the world might come from earthquakes, volcanoes, chemical contamination, fluctuating temperatures or increasing aridity. Forests wither. Embryoes die within the egg. And if that wasn't enough, the falling sea-levels unite previously isolated landmasses, allowing populations of dinosaurs to travel unhindered between the continents. Soon their wanderings bring the added burdens of introduced diseases and new competitors. Most non-avian dinosaur lineages suffer a marked drop in diversity. Across the globe, the number of dinosaur species in any given area can usually be counted on one hand.
In all, an estimated 15-20% of the earth's biodiversity is lost during this time. As their struggle for survival becomes an ever-increasing nightmare, few of the animals notice the new star that has appeared in the heavens. As the weeks progress it gradually grows in size, trailing its gossamer train in its wake. Bigger and brighter it becomes until it becomes clearly visible in the daylight where the volcanic clouds do not mask the view. Then on one night the wanderer seems to lose it's resolve and grows almost imperceptibly dimmer. As the nights pass it's waning continues, smaller and smaller, until it is lost amongst the myriad of stars. The world has arrived at a crossroads and taken one path at the expense of another. For the dinosaurs, the grim struggle to live continues, oblivious to the even grimmer fate that has just been averted.
Gigantala cranitus[]
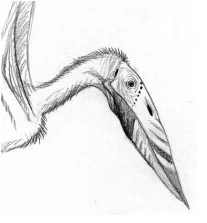
Recovered from late Paleocene rocks in western Canada, the fragmented cranium of Gigantala cranitus represents the youngest traditional pterosaur evidence yet known. A medium-sized azdarachid, Gigantala is in most respects similar to such Cretaceous pterosaurs as Zhejiangopterus and Quetzalcoatlus, although (with an estimated wingspan of 4.5m) not so large. The few other Paleocene pterosaur remnants scattered throughout the northern hemisphere can probably all be attributed the this genus, and as there have yet to be any substantiated pterosaur fossils found from post-Paleocene strata, Gigantala was very likely the last of its kind. However, flightless pterosaurs still exist on the island continent of Australia and nearby areas. Along with that, the fossil record shows evidence of some pterosaurs persisting beyond the Maastrichtian and into the present day. How this was ignored is still anyone's personal guess.
Long ago, it was believed that the great shadows cast by traditional flying pterosaurs soaring high overhead, would be gone forever. The pioneers of vertebrate aviation have surrendered the skies to the birds and recently evolved mammalian bats. Alas, neither of these groups will ever produce a flier that comes even close to rivaling the sheer majesty of the largest pterosaurs... Or so we thought. Upon recent expeditions back to the world Spec, it was revealed that only a few groups of pterosaurs survive in the world of Spec.
LATE PALEOCENE (c.62,000,000 - 57,000,000 b.p.)[]
Depleted ecosystems around the world are regaining their lost biodiversity as conditions improve. Surviving pockets of forest spread outwards across the land, their growth facilitated by the dearth of herbivores. The seas once again teem with plankton allowing marine food chains to reestablish themselves. Insects fill the skies, quickly followed by the birds and early bats. Mammals and lizards scurry in the undergrowth.
Maybe fewer than 100 non-avian dinosaur species worldwide made it through the K-T boundary in sufficient numbers to keep their kind in existence. A roll call of the Northern Hemisphere finds many faces missing, Gone forever are the speedy ornithomimosaurs, the bizarre alvarezsaurs (which still survive in the south) and all but a handful of the horned ceratopsians, both centrosaurs and chasmosaurs. All the northern sauropod species are extinct. Strangely, the bone-headed pachycephalosaurs seem to survive the K-T intact only to vanish before the end of the Paleocene.
Biodiversity remains globally poor, most species are generalists with very large distributions. The runaway volcanism and the horrid black clouds of ash and acid have subsided. However, low sea-levels maintain extensive land-bridges between most continents, keeping regional endemism to a minimum (Europe and North American share more than two thirds of their tetrapod genera at this time) but also giving life the chance to repopulate the most remote of depleted areas.
At this time Africa plus most of the Northern Hemisphere landmasses seem to form one broad biogeographical region. The dominant large plant eaters are hadrosauroid ornithopods, although only three genera are commonly represented a hadrosaurine, a lambeosaurine and, restricted to Africa and Europe, a small basal hadrosaur (Anserodromeus antiquus). Both the giant ceratopsids and sauropods are represented by one genus each, the former found throughout the region, the latter largely restricted to Africa and Europe. Large theropods include at least one tyrannosaur and a moderate-sized abelisaurid. The status of the smaller dinosaurs at this time is unclear - but the presence of hypsilophodonts, leptoceratopsians, small ankylosaurs and maniraptorans is inferred.
The situation in Greater Gondwana (South America, Antarctica and Australia) is patchily understood. The sauropods apparently fared much better in this part of the world with at least 3 genera surviving. Large ornithopods are rare but at least two smaller-bodied families, ancestors of the modern Neodryosauria and Antarctornithopoda, are widespread. The alvarezsaurids are also alive while the ankylosaurs are very rare and scattered. The top predators appear to be giant maniraptoran protobirds although the more primitive abelisaurids are also in force.
As the Paleocene draws to a close, the dinosaurs which made it through the K-T extinction event are well on the road to recovery and diversification. Varying conditions across the globe prod widely distributed species to form clines, then distinct races before ultimately speciating into completely new forms. Their empire may have recently been through hard times but the great beasts have survived and continue to rule the Earth. The Age of the Dinosaurs goes on...
Baropecudis cornuforns []
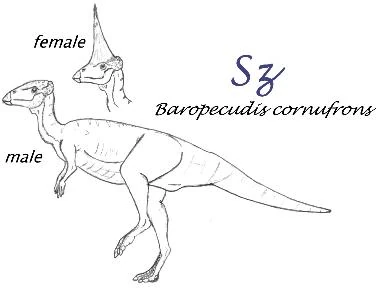
One of the most perplexing puzzles of Specworld's fossil record concerns the disappearance of the pachycephalosaurs. These thick-skulled ornithischians, commonly referred to as "boneheads" or "pachys", first appear in the Barremian of Europe and for the rest of the Cretaceous form a minor component of most terrestrial fossil assemblages in Asia and North America. On Spec, despite having been spared the sudden annihilation of their Arel counterparts at the K-T, the boneheads do not last very far into the Cenozoic.
Baropecudis, informally known as the "Sz", is one of several fascinating fossil forms discovered by Clayton "Reedstilt" Bell's expeditions to Central Asia.This 2 metre long pachycephalosaur, known from several complete skeletons, is notable for the long horn found on some adult skulls. No other bonehead is known to have possessed such a horn although the rest of the anatomy suggests that the Sz (Malaysian for "unicorn") is a derived homalocephalid.
Surprisingly, it seems that only the adult FEMALES possessed this horn, a conclusion based on skeletal morphology and the presence of shell fragments in the abdominal cavity of three horned individuals. Why the females should have such an lethal-looking horn is unclear. Perhaps the females were responsible for anti-predator defence whilst the males fought amongst each other for mating rights with their conventional pachy-heads.
Based on the curvature of the eggshell fragments, it appears that the mother Baropecudis laid only one or two enormous eggs akin to that of an Arel kiwi. The pelvic region of the females was laterally expanded to allowthe passing of such a monstrous object which in life may have been about 25cm at it's longest and 15 cm at its widest.
Importantly, the deposits from which the fossils were excavated are believed to be of either Late Paleocene or Early Eocene in age. If so, this makes it the youngest known pachycephalosaur on Spec. Elsewhere, the boneheads vanish from the fossil record midway through the Paleocene for no apparent reason, a fact made more confusing by the relative abundance of pachycephalosaur fossils in Early Paleocene sites throughout the Northern Hemisphere.
Some Late Maastrichtian deposits (Like the Sandy Site of Arel's South Dakota) would seem to suggest that the pachys were doing quite well despite everything going on around them at the end of the Cretaceous. Early in the Paleocene, boneheads are relatively common throughout the Northern Hemisphere (represented by at least four genera worldwide) and are often the only non-avian dinosaurs represented in some fossil assemblages, the best example being the extinct pachycephalosaur, Spinocephalus superstes. Then, as the Paleocene progressed, something went very wrong. While other small ornithischians began to diversify, the boneheads suddenly vanished, with only Baropecudis surviving till the end of the epoch.
Just what could have happened to bring about the demise of the boneheads in such a short space of time has become a hotly debated topic. The fact that they failed to radiate despite a noticeable lack of competition might suggest that the Paleocene pachycephalosaurs were somehow specialised in ways unknown to us. Their fall would seem to coincide with the diversification of other groups like the protoceratopsians which might have outcompeted them. Or perhaps their breeding habits, assuming that the other pachys laid small clutches like the Sz, left them vulnerable to nest predation. It has even been speculated that their brief success was directly tied in with the conditions of the Late Maastrichtian and earliest Paleocene which so many other species found to be detrimental. When the "bad times" ended they found themselves unable to adapt to the "good times".
The fact of the matter is that we simply do not know what happened to Spec's boneheads.
EARLY TO LATE EOCENE (c.57,000,000 - 40,000,000 b.p.)[]


It is a time of plenty. The continents are altering their outlines and positions as the seafloors spread rapidly. The seas have advanced since the Paleocene, severing land-bridges and facilitating isolation and regional endemism. Global temperatures are warm, nurturing lush tropical floras as far north as Britain. In the seas, warm climate and the return of shallow inland seas put evolution into overdrive. Most modern fish families are now present while the mosasaurs undergo a major diversification event, the archetypal serpentine body shifting into everything from small eel-like river-dwellers to delphinoid pelagic forms.
On land, one could be forgiven for thinking that they were back in the Cretaceous. In North America, the flat-headed hadrosaurine duckbills had branched into a number unique of endemic lineages. But these animals lived in the shadow of the immense Brontoceratops robustus, the most massive ceratopsian to have ever existed. Stalking the herds were the tyrannosaurs which have obviously hit upon a winning formula, being practically unchanged since the Cretaceous. For the most part the chasmosaurs ruled to roost while nearly driving the centrosaur cousins into the point of oblivion.
Across the sea in Eurasia, differences between Old and New World fauna were becoming more pronounced after a long period of uniformity. Hadrosaurines and giant ceratopsids are also present, but were much less common. Here the dominant herbivores were the hollow-crested lambeosaurs followed by the smaller cousins of the ceratopsids, the hog-like leptoceratopsids. The big predators were also tyrannosaurs, but they tended to be smaller and a lot more bizarre-looking than their American cousins. On the subject of Eurasia, the Indian subcontinent collided with Asia about 55 million years ago, eventually giving rise to the Himalayan mountains of today. As for the fauna, a good chunk of it show similarities modern creatures you will find in Spec's Africa, of the Indian subcontinent, it shows that the titanosaurs, abelisaurs, most of deinonychosaurs and possibly ankylosaurs went extinct due to the fact that they couldn't compete with the native fauna in the area. Only the ankylosaurs and deinonychosaurs managed to fare the storm better, but they would eventually succumb to climate change during the Oligocene and Miocene.
Evidence of other coelurosaurs is sporadic, but it is clear that this epoch was a time of diversification for both the oviraptorosaurs and the deinonychosaurs, now the only small predators left in the Northern Hemisphere. Troodontids were more common in the Southern Hemisphere. Mammals were present as small burrowing and climbing insectivores. In Europe and North Africa, there was surprise reappearance of the piscivorous spinosaurs after an absence in the fossil record for 40 million years. Where they had been for all that time is one of Specworld's many unsolved mysteries. In the HE, the spinosaurs went extinct due to climate change and changes in the environment.
Africa, now sliced in half by inland seas, is once again isolated and its dinosaurs are now heading off in their own evolutionary direction. Once again, the land is dominated by herds of giant duckbills, mostly the descendents of Paleocene-Asian immigrants but members of a more primitive local clade are also on the march. Sauropods also flourish as do a host of smaller, poorly understood herbivores. Africa has acquired some Asian tyrannosaurs but they face competition from a much older theropod clan, the neoceratosaurian abelisaurs.
On the other side of the world, this was the final hour for the united Greater Gondwanan dinosaur-fauna, just before the terminal breakup. The last land-connection to South American has only just been severed to Antarctica, while Australia is hanging on by a thread. The sauropods reign supreme; herds of giant long-necked earthshakers are everywhere. Aside from a few duckbills, descendants of a Paleocene interchange with North America, the ornithischians are less conspicuous, but they are still diverse and abundant in the smaller herbivore guilds. Two endemic ornithopod radiations are now recognizable, the medium-sized browsing neodryosaurs and the diminutive omnivorous antarctornithopods . As in Africa, the abelisaurids are on the prowl, but here they play second fiddle to the notoraptors, a group of enigmatic giant sickle-clawed maniraptorans. The status of the dinosaur fauna of India at this time and during the folling period of extinction is presently unknown.
Azolla Event: The Eocene Arctic Ocean
The mid-Eocene was affected by one of the most dramatic decreases in temperature ever witnessed, known as the Azolla Event. Taking place between 49 and 47 million years ago in HE, it resulted in the formation of marine ice caps in both poles for the first time in the Cenozoic, and heralded a wave of progressive climate changes towards the glaciation affected planet of today.
In the Palaeocene, the Arctic Ocean was completly cut off from the remaining seas. The Western Interior Seaway closed completly just a few million years prior, Laramidia was connected with Asia, and Appalachia formed a more or less cohesive landmass with Greenland, uniting thus Laurasia. The only significant seaway was the Turgai Straight, connecting the polar ocean to the Tethys Sea, though in the Eocene the connection between Scandinavia and Greenland was also severed. With the onset of the PETM, this meant that gallons upon gallons of freshwater flooded the Arctic, from newly formed laurasian rivers, increased monsoons and maybe from melting ice caps. So much freshwater in a basically current-less sea lead to the formation of a nepheloid layer composed of freshwater, and maybe subsequent layers of brackish water. If the modern Black Sea is already atypically brackish, the Arctic Ocean was rendered into a massive freshwater lake. How deep these layers were is unknown, but even a layer of a few centimeters was enough to cover the entire sea with freshwater plantlife. One species in particular, the Azolla floating ferns, took advantage of this opportunity, being entirely floating vegetation capable of surviving in the open ocean, far from land. For these plants, the Arctic Ocean was Heaven on earth: 24 hours of Summer daylight, warm water temperatures and a constant suppply of nutrients from rivers. In just a few years, a thick mat would have covered the entire ocean uninterupted, absorbing more carbon dioxide than any rainforest or phytoplankton bloom.
This, of course, led to Paradises's demise: the plants, with their short life spans, would have fallen dead to the seafloor in absolutely gigantic amounts every day, bringing down with them tonnes of greenhouse gas. To make matters worse, the polar nights would effectively kill enitre blooms in a day, bringing to the sea bottom a positively staggering amount of vegetable biomass. Year after year, the planet's atmosphere was cleansed of the massive amounts of CO2 released during the PETM, and, after a period of at most two million years, global temperatures dropped radically. The once fairly warm Arctic Ocean dropped from an average temperature of 13-18ºC (and possibly as much as 22ºC, based on some organisms present in the Eocene) to the current temperature of -9ºC, killing off sea life with the formation of icea caps before salinity levels were even restored. In the span of two million years, the world cooled down to a recognisable temperate climate, though it only became significantly colder in the end of the Eocene, with Antarctica's isolation.
In Spec, the Azolla Event still occured, but it took longer: it's speculated that sea temperatures only lowered significantly 46.7-45 million years ago, and even then the cooling effect was considerably less pronounced, with a minimum of 1-4ºC. It reduced global temperatures, but nowhere near the extreme as in HE, with the start of the severe decline in temperatures posponed to the end of the Eocene, and in fact the Azolla Event's main relevance is in the form of one spectacular evolutionary product of it's existence.
While in HE the *Azolla* ferns were left alone, with no known large herbivore to feed on them, in Spec, as to be expected from a world still retaining it's megafauna, it was different. The fossil sites of Axel Heiberg Island, Denmark and Greenland depict a remarkable marine ecosystem in the Arctic's Eocene, that evolved in response to the reduced levels of salinity.
Only one layer of freshwater was enough to bring Laurasia's freshwater fauna into the Arctic Ocean: in Xel Heiberg, for instance, up to 70% of all present fish belonged to clades previously only known from freshwater. Esociformes radiated into a wide array of species, most notably as pelagic, sardine-like forms that compose up to a third of all actinopterygian fossils. Hybodonts, rare in normal marine biomes, became the dominant sharks, while gars and paddlefish expanded at the expense of ichthyodectids and pachycormids respectively. Truly marine fish compose as little as 10% of all fish, all of them benthic, even even then most benthic niches were taken by brackish water forms like sturgeons and rays. Freshwater insects are also present, while known crustaceans don't seem very different from other marine ecosystems. Macrobaenid turtles, alligatorids, Borealosuchus and the first marine neochoristoderes are also known from both Axel Heiberg and Greenland, as are dermatochelids, the polycotylids Oceanocursor and Njordia, a possible mosasaur, the hesperornithes Arctophoebe, Freyavis and Okeanides, ebergs, the possible petrel/loon/penguin relative Alcagavia, the ichthyornithe Vipelarus and the ornithocheiroid pterosaurs Phoenicorhamphus and Boreodraco.
The Azolla Event did more than just colonise the Arctic with freshwater fauna, however. The huge mats of floating vegetation were without doubt a bait to herbivorous dinosaurs, which would already be common around the Arctic Ocean, searching for seaweed or bathing to relieve the heat. In testament of these, hadrosaur fossils are almost omnipresent in all fossil sites pertaining to the Eocene of the coastoal Arctic, hundreds upon hundreds of animals seemingly marching across the polar shallow waters throughout the Azolla Event, wading like giant dinosaurian hippos. A few genera took this further, adapting to a more aquatic lifestyle to take advantage of the initially hadrosaur-untouched mats in open waters, and one, Pelagoanatosaurus, has become the only known fully aquatic hadrosauroid, it's hands becoming flippers.
However, hadrosaurs weren't the only dinosaurs exploiting Azolla blooms. Another clade of unlikely explorers moved into the ocean, and unlike the duckbills, they managed to keep their exploration of aquatic niches, surviving to this day as dinosaurian pioneers.
These were the titanosaurs, the lineage known as Mokelosauria.
Unlike hadrosaurs, common in humid biomes, titanosaurs were for the most part rare in humid biomes, being primarily dry-highland specialists. Indeed, looking at titanosaurian collumnar limbs and fingerless, almost horseshoe like hands, it's clear why these dinosaurs were rare in areas with soft substrates. So the evolution of the bizarre mokeles had been one of Spec's greatest mysteries. Thankfully, the fossils of the Eocene of Denmark gives us a clue on how these sauropods might have become aquatic.
During the PETM, titanosaurs in Laurasia saw themselves in quite the predicament, as the tropical rainforests, even if not as dense as in HE, posed something of a challenge to these open-space dinosaurs. Most were forced northwards, into the colder polar forests, but some simply moved to the coasts, exploiting the open habitats in beachlines, especially in Europe where migration was severely restricted. Living in coastoal areas meant that sauropods probably always have foraged for seaweed, but it was during the unique conditions of the Eocene that marine plants bloomed, with the appearence of seagrass.
One particular european lineage was in the right place at the right time: in Scandinavia, beachdwelling titanosaurs, seemingly related to older european island endemics like Magyarosaurus, benefitted from the blooms of Azolla, that quickly covered chunks of the Arctic. Before even hadrosaurs moved into the sea, sauropods were already wading after the ferns in shallow waters, their boyancy aiding them in foraging on such a pelagic banquet. Over time, the horseshoe like hand configuration changed into a flatter profile, while the metacarpals widdened, allowing the animals to propel themselves more easily, and while not fully efficient to support weight on soft substrates - the the more elephantine hindfeet make up for that, though -, it at least allowed them to become very efficient swimmers and bottom waders. One of the earliest known recognisable mokeles, Arctomokelosaurus, exemplifies this hypothesis: the first species, A. eonatans, is a fairly typical sauropod, only differing significantly in wider metacarpals, but posterior species like A. antiquus and A. litoralis show a progressive flatenning of the "hand-horseshoe", and this it taken to the logical extreme in A. pontus, where the forelimb is almost flipper-like.
Thus, in the mid-Eocene, herds of floating titanosaurs cruised the Arctic, feeding on the open water mats of Azolla while most hadrosaurs remained in the shallows. By the end of the Azolla Event, another lineage of aquatic herbivores joined them, the duckgongs. Relatives of the fast jackalopes and slow vanguards, duckgongs are presumed descendents of animals like Thescelosaurus, already suspected of semi-aquatic habits, and they first appear in the Green River Formation. They quickly spread into the sea, taking advantage of budding seagrasses, and their late appearence in the high Arctic is presumed to be largely due to the isolation of the sea. The local two genera, Actoanser and Barotarsus, however, seem quite basal among duckgongs, whch has led some experts to believe that they instead descend from north american freshwater endemics.
All these herbivores left a tremendous impact in the ecosystem of the Arctic Ocean. One of the two local polycotylids, *Njordia*, has developed raptorial features akin to those of long gone pliosaurs, and combined with fossils of dismembered hadrosaurs, it is thought to have preyed on these aquatic herbivores. Likewise, the simoedosaurid *Anguillasuchus* also has deeper jaws with serrated teeth, one of which found in a mokele neck vertebrae. Early mokeles indeed already have a full scute armour like their modern relatives.
A more important change, however, occured due to the activity of these animals. Clearing the sea surface of Azolla has allowed for more sunlight to reach the seabottom, allowing for the development of other aquatic plants. The ocean floor would have been covered by various types of flora depending on the water conditions, with more familiar freshwater angiosperms in shallower, less aline water while kelp and seagrass bloomed on water with a higher amount of salt content. These underwater plains anf forests would also have been aided by the enormous amount of dinosaur guano, breaking down competing Azolla into nutrients. Mokeles once again seemed to have been the pionners, starting to lose their bone pneumacy early on. By the end of the Azolla Event period at 46/5 million years ago, the Azolla mats began to loose relevance in favour of sea bottom plants in terms of CO2 containment.
As delayed as the Azolla Event was, temperatures eventually did decrease, and while full glaciation in the Arctic didn't occur, sea temperatures became too cold for most endemic life forms. Precipitation in turn decreased, and the extension of the Atlantic into the Arctic ended the isolation of the sea, allowing currents to set in. The freshwater biome was thus destroyed, the mats of Azolla and other freshwater plants disappeared, and so did most of the sea's endemic fauna, either due to the end of suitable conditions or due to the invasion of atlantic marine animals, especially mosasaurs. The herds of marine hadrosaurs, deprived of food and subjected to new marine predators, were utterly decimated, with the only marine ornithischians for the rest of the Cenozoic being duckgongs.
Mokeles, however, survived. Fossils of mokelosaurs outside of the Arctic Circle are known soon after their appearence, with Turgaia and Ferroserpens known from Kazakhstan and Europe, having swam down the Turgai Straight. A few authorithies have also suggested that true mokeles evolved in the european Tethys from island endemics and that the Arctic species are an altogether different clade of sauropods, but evidence so far tends to the Arctic hypocthesis, as european mokeles don't have the same sort of adaptation ladders, being already primarily aquatic animals.
Whatever the case, mokeles proved to be quite successful, expanding in the opposite direction of that of HE's tethytheres, colonising Europe, Asia and Africa from the north. They expanded along the vast seagrass plains of the Tethys, but they quickly explored freshwater biomes, learning to return to a browsing existence, now free from the contraints of dense forests and soft soils, simply foraging from the water on riverine trees and plants, though eventually some returned to a terrestrial existence. By the Oligocene, mokeles were everywhere but Australia and South America - though some fossils from the Oligocene and Miocene may argue otherwise -, and even though marine species eventually all but vanished, replaced by duckgongs, mokeles are still going strong on the freshwater bodies in most continents.
One world's calamity is another's way of creating an inovative design.
Balaenanguis primitivus[]
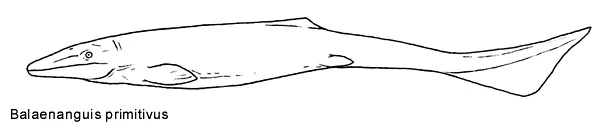
Recently-discovered of Balaenanguis, the most primitive known lizardwhale which grew up to lengths of five meters, from early Eocene sediments in Europe, sheds much light onto saurocete evolution, especially the lineage of the eninigmatic Nile mosasaur, genus Apep.
Eohadrus excelsior[]
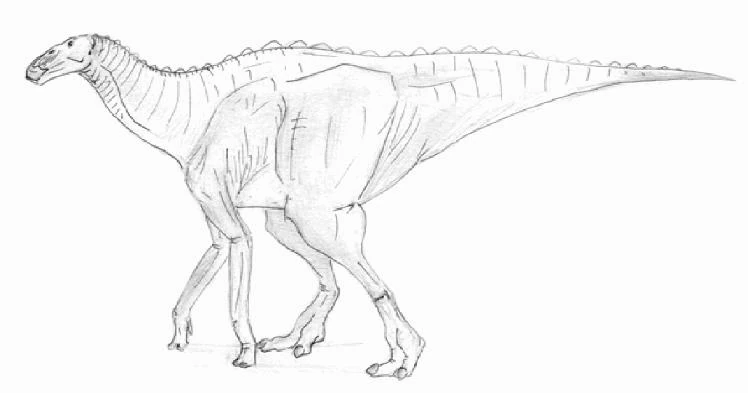

Unearthed in from 50-million-year-old Eocene deposits of Wyoming, ten-meter-long Eohadrus excelsior is one of the best-preserved specimens of hadrosaurid diversity during the Cenozoic. The skull morphology of this species is quite similar to that of its Cretaceous ancestors such as Kritosaurus and Bactrosaurus, with a blunt, toothless beak and layers of small, diamond- shaped teeth in the back of its mouth. The limbs of Eohadrus, however, are quite distinct from those of its ancestors, the forelimbs being roughly the same length as the hindlimbs. Eohadrus, then, was a fully quadrupedal galloper, like modern hadrosaurids, and unlike the still partially bipedal basal hadrosaurids of the Mesozoic. Eohadrus's neck was long and flexible, and its tail, no longer necessary for balance, but still the anchor for the muscles of the legs, was deep and blunt. Unlike its plains-dwelling descendants, Eohadrus was most likely a forest-living animal, browsing from the ground and from low-growing trees.
Megacaudia nameni[]
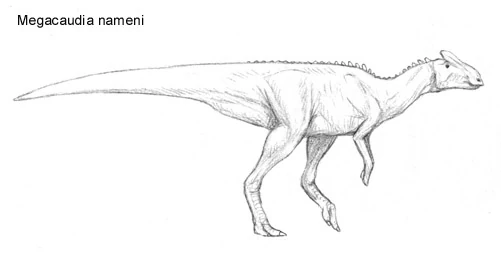
Growing up to lengths of 5.5 meters longs and weight up to half a ton, Megacaudia nameni was an early member of megacaudinae living in the late Eocene North America. These fleet-footed hadrosaurids already had many distinctive characters of the later megacaudines, though in many respects they were more primitive than some other species uncovered from the same period. Despite its weight, this creature was a surprisingly agile creature and most likely fulfilled a similar niche to the extinct ornithomimids.
Brontoceratops robustus[]
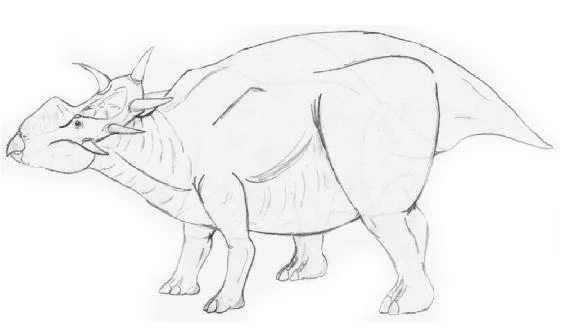
Brontoceratops robustus, the Eocene Chasmosaurine, was the largest marginocephalian ever. While its length (twelve meters) has been surpassed by some Eurasian ceratopsians, its weight (ten tonnes) is unrivaled. These bulky creatures swaggered across the American interior in thunderous herds. Males had a ridge over their shoulders that may have supported a fatty hump that cushioned their frills, reducing the stress applied to them when two huge bulls collided.
Gigantops dominus[]
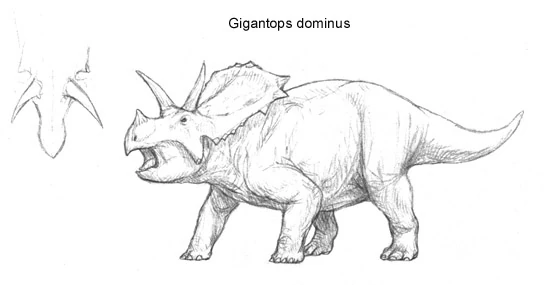
The Eocene was the last, yet most glorious chapter in the history of chasmosaurines. While North America was ruled by the massive Brontoceratops, Eurasia was a home to another ceratopsian giant. The chasmosaurine ancestors of Gigantops had crossed the land bridge between Asia and North America during the early Eocene and at the time ruled all the northern continents. Gigantops was quite likely the largest of the Eurasian chasmosaurines, growing up to nine meters long, with a head as long as that of Torosaurus, even though the neck frill of Gigantops was proportionally shorter.
Eobrachioceratops antiquus[]
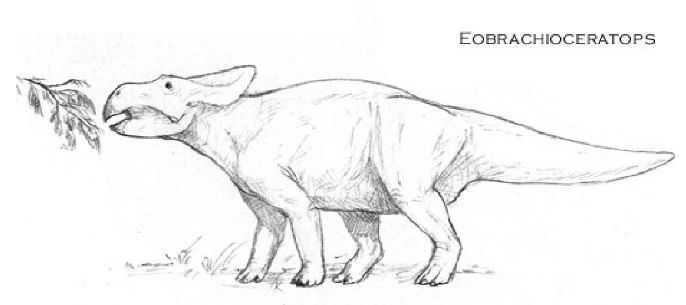

The Eocene Eobrachioceratops antiquus is the most basal brachioceratopsian yet known. Though this three-meter-long animal wasn't much different from the other leptoceratopsian forms of that time, its forelimbs were already noticeably longer than the average.
Spinotitan grandis []
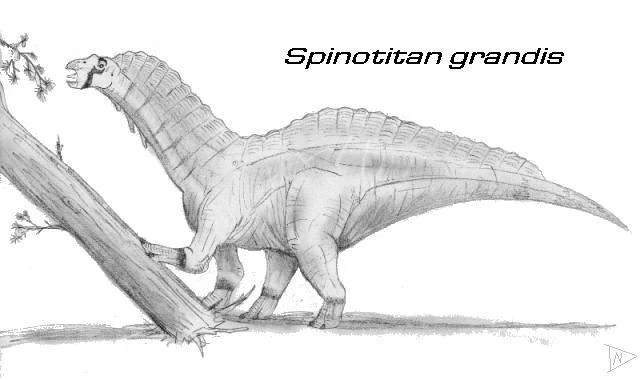
Looks can be deceiving and in the case of Spinotitan grandis, it applies perfectly. When it was first discovered, it was once believed to be an aberrant species of late-surviving spinosaur, in South America to be precise. However, later expeditions revealed that this was actually a species of medium sized titanosaur; growing up to 60 feet long and weighed up to 10 tons. Not only was this a magnificent case of convergent evolution, resembling the high-spined sauropod Amargasaurus, but also case of mistaken identity.
Piscisraptor nauticus[]
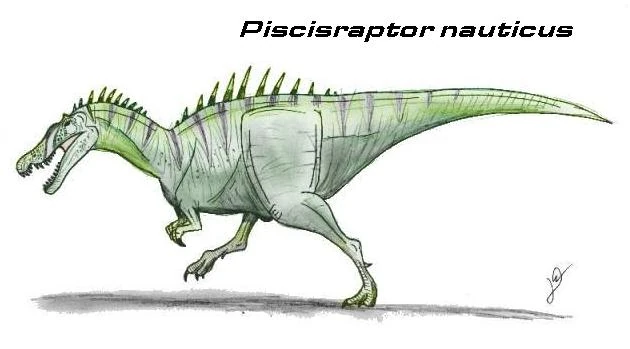
Piscisraptor nauticus was a medium sized spinosaur from Northern Africa, growing up to nine meters long. Fish bones found near its stomach cavity suggest it was a piscivore like its ancient cousins. It is believed that Piscisraptor was driven to extinction by the faster and more powerful abelisauroids.
Carnosuchus tarbos []

Carnosuchus tarbos, colloquially known as "The Big One", was last of the afrosuchine spinosaurs, some of the last spinosaurs in general, and the last spinosaur overall which grew up to lengths of 11 meters long. Adapted for hunting other dinosaurs rather than fish, it possesses a larger, stronger skull and raised neural spines much like the long-extinct Acrocanthosaurus. It went extinct when the smaller priscataurs out competed it for food. With the extinction of Carnosuchus tarbos, the history of the spinosaurs finally came to an end.
Phobotyrannus robustus[]
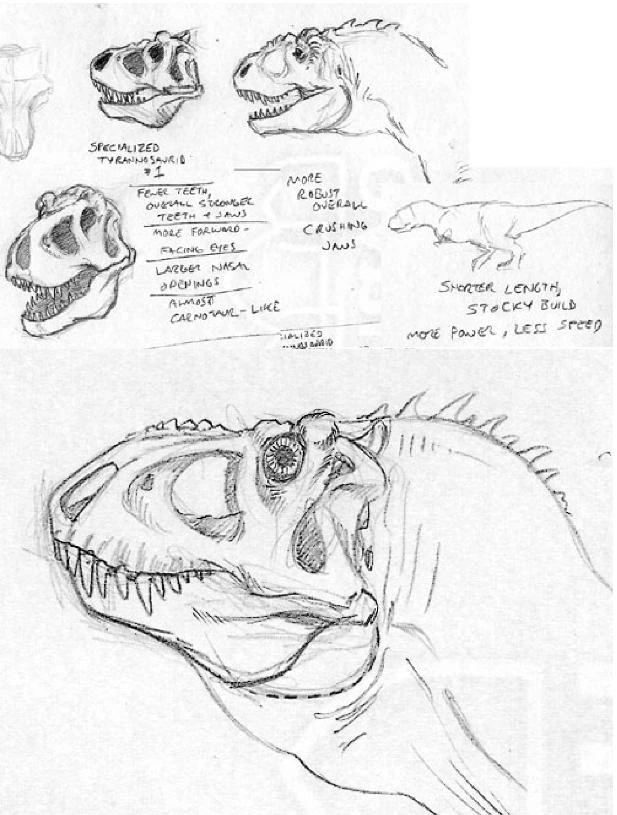

Phobotyrannus robustus was a large, growing to 10 to 12 meters long, powerfully built ceratotyrannid, an abberant branch of Tyrannosauroidea that appeared some time in the early Eocene and then went extinct in the Miocene. This creature was found in fossil deposits through out Asia, most notably in Mongolia and China, where the fossil remains of Eobrachioceratops and Gigantops were unearthed as well. Implying that its primary prey was mainly ceratopsians. This hypothesis is further backed up with evidence from the fossil records; a recent skeleton of Gigantops dominus was revealed to have bite marks from teeth matching those of Phobotyrannus robustus.
Orgetorix ozymandias[]
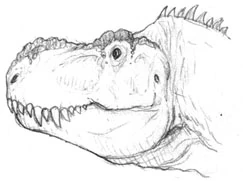
A late Eocene ceratotyrannid from Europe, Orgetorix ozymandias is known from a partial skull and limb material, and is estimated to have been close to 15 m in length. Like its relatives, this massive carnivore had robust skull a possibly a bone-crushing bite. With no ceratopsians in the area, Orgetorix most likely sustained itself on the large number of lambeosaurs and hadrosaurs in the area.
Arbroraptor mongoliensis[]
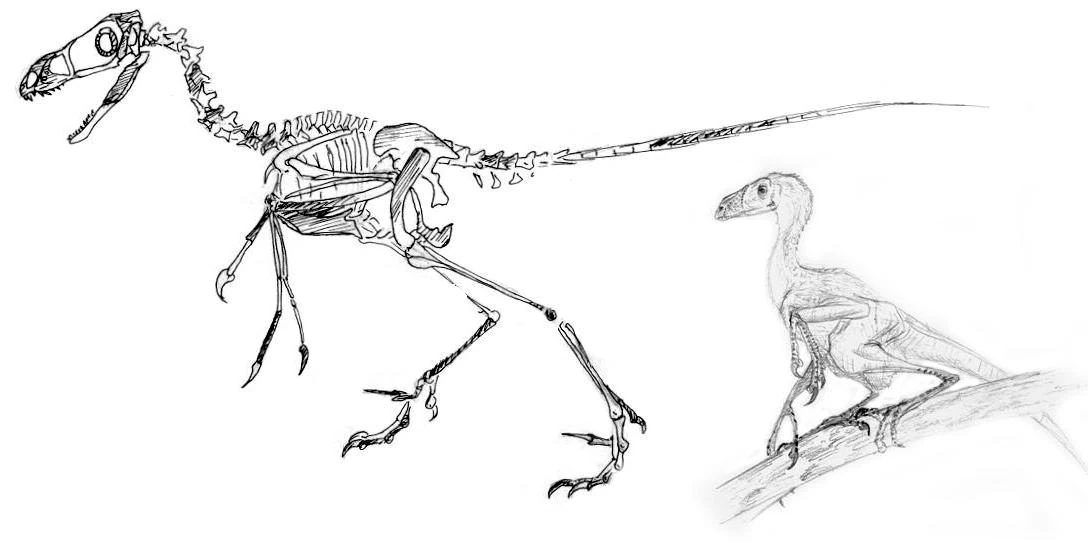
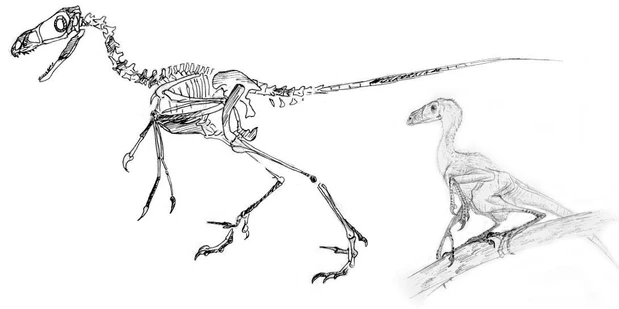
Found in the Eocene deposites of Mongolia, the meter long Arbroraptor mongoliensis is the earliest known undisputed arbronychosaurid. Remains of this species have been found across central Asia, and A. mongoliensis has shed a great light upon arbronychosaurid evolution. Apparently, by the Eocene, arbronychosaurids were already well established as climbers. A. mongoliensis sported fully reversed hallux claws on its feet as well as thumbs that opposed the other two fingers in its hands. The hyperextendable inner toes, a distinguishing feature of the deinonychosaur clade (of which arbronychosauridae is a part) were reversed, but had yet to come into full opposition with the two front toes of the foot. It is believed, therefor, that A. mongoliensis was not as surefooted in the trees as its descendents.
Palaeocarpo primitivus[]
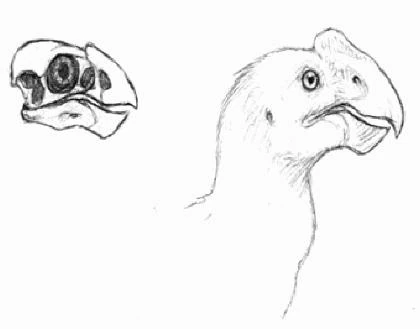

Palaeocarpo primitivus is one of the best known primitive pithecaviforms, dating back to late Eocene. These crow-sized flightless birds have been found in all the northern continents, and probably mainly lived on fruits and nuts. Despite being named after the crestless carpos, several Palaeocarpo species had crests on their head. Being a basal pithecavian, it was nevertheless quite closely related to the forms from which the modern carpos descend.
Patriopokemus mordax[]
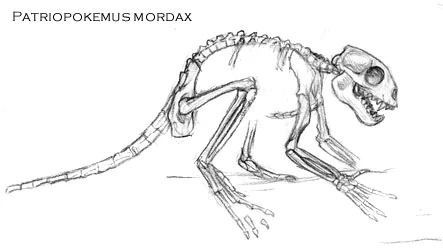
Patriopokemus mordax is considered to be the common ancestor of all Pokemusidae, even Parapokemus. This Eocene proto-primate was only 25 cm long and was probably an omnivore or insectivore. Depicted here are is the most complete Patriopokemus specimen, which is missing only a few bones besides the ill-preserved ribcage.
Gruorostrum tingamarri[]
An Australian theropod, most likely a species of deinonychosauria, from the Eocene. It seems to be a basal maniraptoriforme, with a few therizinosaur like features like a weight supporting first toe and a beaked snout (with no teeth though); with blunted claws like oviraptors it seems to had been more carnivorous than their northern look alikes, most likely preying on aquatic invertebrates like molluscs and crustaceans.
EXTINCTIONS AT THE EOCENE-OLIGOCENE TRANSITION (c.40,000,000 - 31,000,000 b.p.)[]


During the closing phase of the Eocene Epoch (the Priabonian) and the opening phase of the Oligocene (the Rupelian), dinosaur communities worldwide were once again plunged into crisis. Across the globe, countless species were facing extinction, both on the land and in the oceans. The direction of evolution on the Specworld was to be forever altered.
The extinctions of Eocene-Oligocene transition did not take place as a single discrete event, but rather as a series of extinction pulses. The first took place close to the start of the Priabonian about 40 million years ago with three or four additional pulses taking place throughout the Eocene-Oligocene transition, the last one occurring 31 million years ago towards the end of the Rupelian. A similar phenomenon occurs in Arel's fossil record and the progression of events in both timelines show strong similarities with regards to the marine fossil record. In the oceans during the extinction pulses, those groups most seriously affected were generally tropical and inshore forms. Many foraminiferans disappear, and those that survive show a heightened amount of oxygen 18 in their tests (a phenomenon associated with a lowering in temperature). Tropical calcareous nanoplankton are decimated whilst high-latitude floras of these organisms appear at the equator.
The evidence points to both a cooling of the world's oceans as well as a severe drop in global sea-levels during the final pulse of extinctions in the Oligocene. Such conditions at sea would in turn have a direct consequence for life on land. Although there is some evidence of extraterrestrial impacts during this time, it seems likely that this climatic deterioration and fluctuation was tied in with final breakup of the ancient Gondwana.
Towards the end of the Eocene, Australia rifted away from its last connections with Antarctica. Cold water flowed into the widening gap, deflecting the warm currents that had previously kept the south polar climates mild. For the first time since the Paleozoic, a permanent ice cap began to form at the South Pole. With the formation of the psychrosphere, a great chilling engine was created in the southern oceans. Warm water was trapped in the south where it was cooled and forced to descend into the ocean depths before spreading back towards the equator. Each time the cool temperatures were sent towards the lower latitudes via wind, oceanic currents or upwellings, it triggered off a wave of extinctions. In the longer term, this marked the start of a profound shift in the Earth's climatic history would ultimately lead to the ice ages of the Pleistocene.
The exact timing of many of the terrestrial extinctions is unclear. Most dinosaurs seemed relatively unaffected by the first extinction pulse at the the start of the Priabonian (40 million years ago), but took losses close to the Eocene-Oligocene boundary. Overall, the climate on the land seems to have gotten colder and drier during the pulses. As seas retreated, once coastal regions were left far inland and faced increasingly dry conditions. Areas once blanketed by humid forest gave way to semiarid scrub, and for the first time, grasslands.
For Brontoceratops and other horned giants, this was their final curtain call. The great evolutionary dynasty of the chasmosaurs had come to an end. The hadrosaurs had faced hardships as well. The lambeosaurs, which had been most adapted to moist woodlands and swamps, suffered large casualties. The few reaming species of Eurasian saurolophinae had fared better.
In North America, however, things were looking much brighter for the hadrosaurs, most notably the saurolophinae. In fact, the early Oligocene was time of incredible diversification for the American hadrosaurs. By the end of the Oligocene, many hadrosaurs, from massive grazers to tiny antelope-like bipeds, could be seen across all over North America.
Africa was also experiencing a duckbill bloom. Although losing most of its larger, Eurasian-derived species, the more primitive endemic hadrosaur clade was holding the fort. One lineage began to develop an unusual hoof-like hand and was destined for greatness later in the Cenozoic.
Across the Northern Hemisphere and Africa, theropod extinctions appeared to have been fairly minor and localized. Most seem to have adapted to hunting new prey species with little difficulty. The only significant loss were the spinosaurs, vanishing after their brief reappearance, this time for good. The herbivorous therizinosaurs, some of them already adapted to semiarid conditions, increased their diversity during this time.
Being closer to the spreading coldness that is the root of this calamity, life on the fragments of southern Gondwana was in complete disarray. The great browsing sauropods were in dire trouble and were soon reduced to scattered remnant populations. By the early Oligocene, they had vanished from Australia and Antarctica while just barely being able to cling on in Africa. In South America, the lone genus Acrotitan survived the crises. The South American duckbills also disappeared, confusingly as they were closely related to forms that did well in North America. Perhaps their life-cycle was somehow tied in with the doomed sauropods? We simply don't know.
The combination of cooling climate and prey loss lead to the almost total annihilation of the large Gondwanan predators. In Antarctica and Australia, every theropod species bigger than 100kg failed to make it into the Late Oligocene. Like the sauropods, the abelisaurids found temporary refuge in South America, surviving there long enough to be obliterated in the Pliocene by North American invaders.
The only dinosaurs that seemed to be holding their own were the small bipeds. Polar populations of neodryosaurs and antarctornithopods retreated northwards from the spreading ice and found large swathes of Australia and South America suitably habitable. With so many of their predators and competitors missing, they quickly make themselves at home.
A brief look at the oceans shows many changes there as well. Shadowing the loss of the archaeocetaceans of Arel, the ancestral long-bodied, four-flippered mosasaurs were in trouble, surviving only as a few diminuative freshwater forms. However their more derived torpedo-shaped pelagic kin, the recently evolved saurocetes, were doing fine. Surprisingly the ancient ammonites had been given a new lease on life. Reduced to a handful of cool-water species since the Maastrichtian, the disruption of equatorial marine ecosystems gave them a crucial window of opportunity to spread and diversify.
Unlike the Selandian extinction event, the Late Eocene crisis saw the loss of relatively few family and order sized groups. Certainly, the world biota after the Late Oligocene stabilization was in no way impoverished. In fact, a number of clades seem to have increased in both population and diversity. The Eocene-Oligocene boundary, then, was less a time of catastrophe and more a period of high species turnover, with new forms quickly stepping in to replace the old.
SPEC'S LATE OLIGOCENE (c. 30,000,000 - 24,000,000 b.p.)[]
After millions of years of climatic seesawing, some semblance of stability had at last come to the world. The fragments of Gondwana were now fully separated. Australia and India crawled northwards, while Antarctica, now isolated in the extreme south, continued towards increasingly frigidity. The climate still supported vast tracts of lush greenery, but now there was a distinct hint of drier times to come in many parts of the world. Here, the forests were giving way to savanna woodlands where the recently evolved grasses were playing an increasingly prominent role.
In many ways, the dinosaur fauna of the Oligocene was the most bizarre of the entire Cenozoic. It was a world that mixed the last survivors of relict clades with early representatives of familiar present-day families. Many of the old-timers were giants including the Asian lambeosaur Sauropodimimus and the South American sauropod Acrotitan. However, these old survivors were outnumbered by a host of weird and wonderful newer forms.
The Oligocene was the peak of leptoceratopsian diversity, with the clade represented in Eurasia and North America by everything from tiny forest runners to titanic long-necked browsers. Duckbills of all shapes and sizes marched across North America and Africa, although they were not very diverse in Eurasia. Here, the most common ornithopods were the eurolophs, enigmatic creatures with no close living relatives, except for the strek and a few others. Strange herbivores, unrelated to anything elsewhere, also appeared South America and Australia.
The carnivores seemed to also get into the spirit of weirdness. Northern tyrannosaurs sported all manner of strange cranial crests, as did the abelisauroids of South America and Africa. In Australia, flightless pterosaurs and a group of ornithopods called rhynchoraptors seized the top predatory niches, breaking the theropod-monopoly for the first time since the Triassic.
In terms of family-level diversity, most researchers consider the Late Oligocene to mark the zenith of the Cenozoic dinosaur fauna. As the Oligocene drew to a close and the Miocene began, however, this diversity began to wane. In the oceans and throughout most of the southern landmasses, it was business as usual (aside from unusually high planktonic turnover). However, in a greater part of the northern hemisphere, the dinosaurs were vanishing, losing close to one third of their species diversity.
Procurvihadrus specensis[]
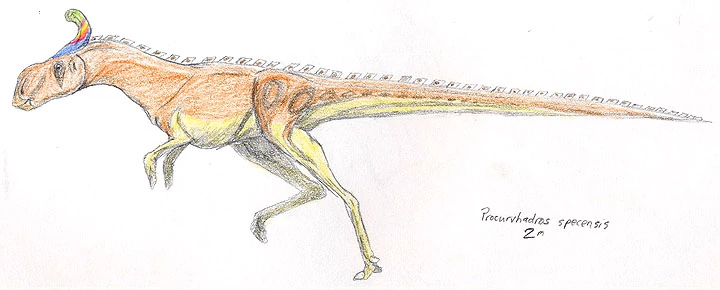
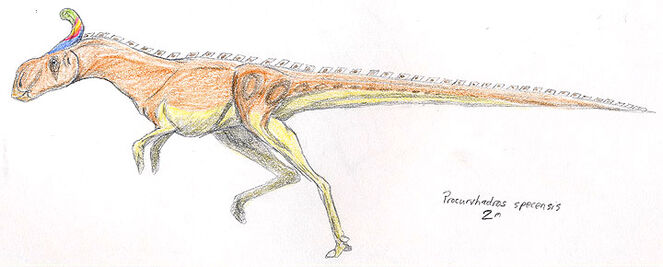
Growing up to two meters in length, Procurvihadrus specensis is the best known member of the megacaudinae clade and illustrates most of its kin's salient qualities. The head is large and boxlike, the forelimbs are reduced and the fifth and first digits are lost, leaving the rest to form a hoof-like pad. The hind legs are long and slender, with a pronouncedly digigrade. The most strangest part of this animal's anatomy (and the namesake of the clade), the tail, is long and rodlike, stiffened with a network of ossified tendons to form a balancing appendage quite similar to that of a deinonychosaur.
Paramegahadrus vetus[]
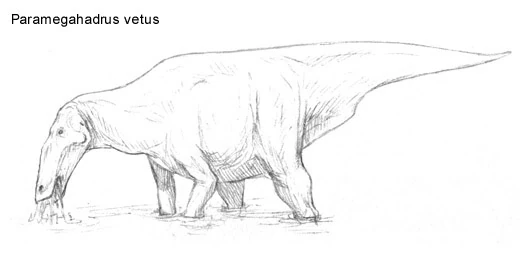
Paramegahadrinae is an extinct group of North American hadrosaurs that appeared in the middle Oligocene. They were first thought to have been the ancestors of megahadrines, but certain details of their anatomy, most importantly the shape of the head and the shortness of the forelimbs have lead paleontologists to believe they were actually a primitive evolutionary offshoot of Hadrosauroidae. It seems that while megahadrines eventually became grazers of the prairies, paramegahadrines remained in the marshlands and riversides where their common ancestors had evolved. It is still unclear what events exactly lead into their demise, however, the most likely conclusion being climate change. Two of the most noticeable examples being Paramegahadrus vetus and Megabeluasaurus magnacephalus. Growing up to lengths of 8 meters long and weighing 2 tons, Paramegahadrus vetus remains have been unearthed in North America near ancient swamp deposits, from Mexico to Oregon.
Protobrachioceratops gracilis[]
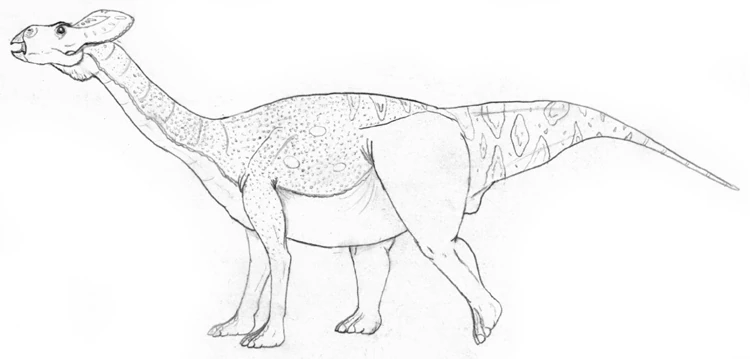
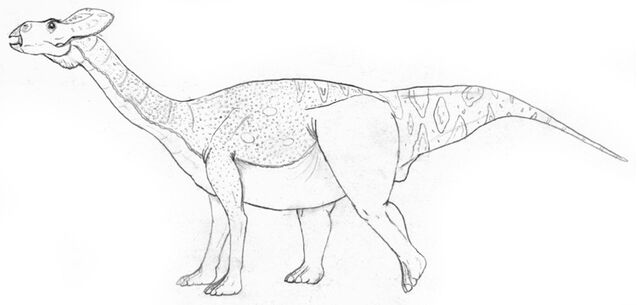
At the height of leptoceratopsid evolution, 'Sauroceratops' (Protobrachioceratops gracilis) was the largest of the clade. This Oligocene browser has evolved convergent with sauropods and its closer (though still distant) relatives, South America's false-sauropods. 'Sauroceratops' measured 14 meters, but since most of its length was taken up by its long neck and tail, it weighed surprising little for its size, only 6-8 tonnes. The first half of its tail was slender, yet very deep. It was probably brightly colored to attract mates or intimidate rivals. The frill, prominent in other ceratopsians, has been reduced, making it easier for this high-browser to feed. It should be noted that this was originally named "Titanoceratops gracilis", but this was changed in order to not confuse it with the massive chasmosaurine which was discovered back in 2011.
Errosaurus borealis[]
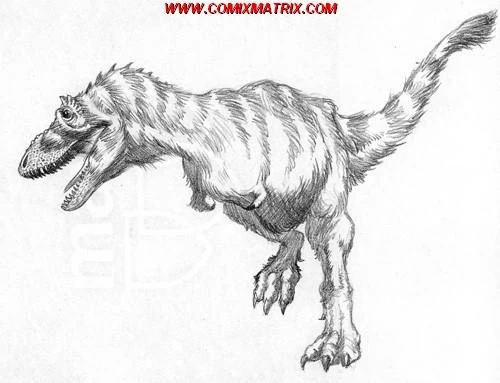
As seen in the picture above is a reconstruction of Errosaurus borealis, an Oligocene species unearthed in western Canada. E. borealis is the earliest known North American errosaurid. The presence of this dinosaur on the North American continent reveals how quickly clade Errosauridae spread from its origins in Eocene Eurasia. On a similar note, fossils show that this creature grew up to lengths 5-6 meters long. A stark contrast when given the fact that most species of Errosaurus grow to lengths only a few meters long.
Bavarionyx cursor[]
If paleontology serves a purpose, it is to remind us, again and again, that the current breadth of life on Earth is but a meager fraction of its former diversity. We have all, by now, become accustomed to the fleet-footed yales, the lumbering hmungos, the sumo-esque therizinosaurs, but as little as two million years ago there existed a group of herbivores completely unlike any seen today.

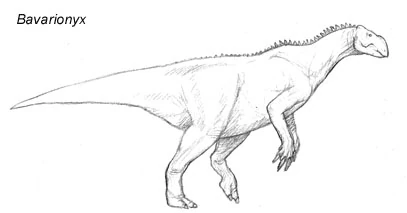
The eurolophs (order Eurolophia) appeared suddenly in Europe, fully diversified into a number of distinct forms, about 30 million years ago, ruled the continent through the Oligocene, and then disappeared at the end of the Pliocene. Although eurolophs are clearly ornithopods of some kind or other, the eurolophs do not fit in with any other branch of this tree. Their cranial anatomy is advanced, with a long muzzle and three rows of cheek teeth to form a "dental battery" like that of a hadrosaur. However, the jugals (cheek bones) tend to be pronounced, as in the basal ornithopods like Heterodontosaurus, and the ossified tendons that stiffened the tails and spines of every ornithopod since the Jurassic are completely absent. As of today, the larger species of Euroloph are long gone, however, smaller species of the Euroloph, most notable the struthiopods (which not to be confused with the jackalopes) stil survive to this very day.
From Oligocene Europe, Bavarionyx cursor is the earliest relatively complete euroloph yet discovered. Growing up to the same size as an ostrich, this creature is supposed to be an early rhinolophosaur, but shares some features (including its bipedal stance) with the struthiopods. Such a "missing link" may support a european origin for the eurolophs, but the evidence is still fragmentary. This is one of the most controversial fossils found in the world of Spec. Bavarionyx cursor, a bizarre bipedal species of Eurolope that went extinct before the start of the Miocene epoch.
Bellasaurus leidyi[]
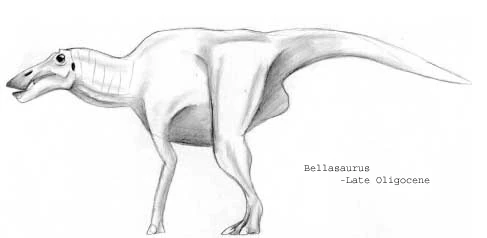
Of the three great hadrosaur groups of the Oligocene, the hippohadridae, not to be confused with modern day hipposaurinaes of the neohadrosauria which inhabit the Americas and Eurasia and the hipposauridae therapsids which inhabited South Africa during the Permain period, were undoubtedly the most diverse and arguably the most successful. While the paramegahadrines were giants and the megacaudidaes were probably more numerous, hippohadridae occupied a much wider range of forms, from tiny, hoglike browsers to giant grazers rather like modern hmungos. Growing up to lengths of 20 feet long, Bellasaurus leidyi is one such example.
Like the megacaudine hadrosaurs, hipposauridaes retained only the middle three digits of their forelimbs, but these sported large nails that probably extended past the hoof-pad. Most sported enlarged neural crests over the shoulders to anchor powerful grazers' neck muscles, and many later species possessed a very deep lower jaw, perhaps to aid in cropping early grasses. Hippohadridae tails tended to be rather narrow and stubby, possibly because their function as balancing devises were irrelevant to the quadripedal herbivores.
Pygmihadrus mirus[]
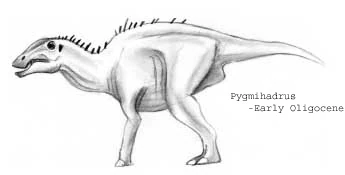
Hippohadridae, not to be confused with the modern day hipposaurinae or the exinct hipposauridae therapsids, probably evolved some time during the Eocene, but their fossil record begins in the early Oligocene with such genera as Pygmihadrus, a meter-long herbivore with short, stumpy legs and tail. Fossils similar to this one appear throughout the Oligocene and Miocene.
Gryphorhynchus robustus[]
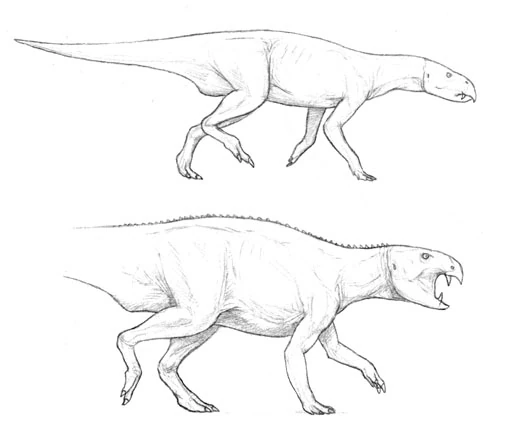
Above: the old restoration of Gryphorhynchus robustus (Australia) shows it as a sleek predator, not unlike modern rhynchoraptor. Below: The new more accurate restoration. The tail of the animal is still unknown.
New fossils unearthed from Late Oligocene deposits at Spec-Riversleigh have proven that the two meter long Gryphorynchus, once thought to have been the first rhynchoraptoran predator, to be something else instead. Until now Gryphorhynchus was known from an incomplete skull and some fragmentary limb and pelvic material. Based on the fossils scientists had reconstructed the animal as a sleek and swift predator not unlike the smallest modern rhynchoraptors. It was thought to be closely related to the common ancestor of rhynchoraptors.
The newly found Gryphorhynchus skeleton, nicknamed "Greg", has proven these reconstructions flawed. The new animal that belongs to a new species, Gryphorhynchus robustus, that was clearly closely related to the earlier fossils. In fact it is possible that the bones found earlier belonged to a juvenile G. robustus. Scientists estimate this creature grew up to lengths of two meters long.
"In life Greg was a big and bulky beast and probably not much of a runner. It's more likely that he was an omnivorous scavenger," said Specpaleontologist Kim Xu, who was involved in the excavation of the animal.
Though the otherwise nearly complete skeleton is missing nearly all of the tail, it has become clear that Gryphorynchus was too different from Rhynchoraptors to be a closely related to them. "It likely now that the antarctornithopods that gave rise to rhynchoraptors experimented with omnivory and maybe even carnivory several times," Xu said.
"This means the earliest rhynchoraptoran remains are now from the Miocene," said professor Brian Choo, who has been studying fossil rhynchoraptors for years. "But that doesn't necessarily mean they aren't there. We just haven't found them yet," he added. From Spexploration Weekly, November 2003 Issue. - Brian Choo and Daniel Bensen
Gryphorhynchus aenigmaticus[]
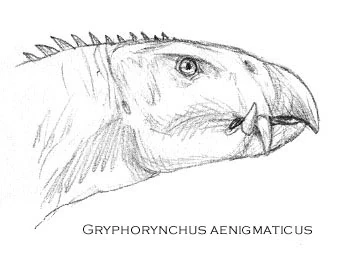
Gryphorhynchus aenigmaticus was a 2 meter long early rhynchoraptoran, though appearing much later after Gryphorhynchus robusuts, known from a nearly complete skull and fragmentary limb material. It has been suggested that this species was a scavenger, but other paleontologists postulate a lifestyle of active hunting. This is a rather controversial specimen given the information that was gathered about extinct rhynchoraptor back in 2003. More research should settle this issue some time in the near future.
Undescribed Insect-Eating Therizinosaur[]
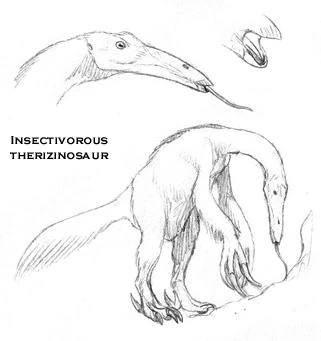
Early paleontologists excivating in Spec's eastern Eurasiawere quite surprised to discover the scattered remains of what appeared to be a group of tiny, ant-eating therizinosaurs, with elongated tubular snouts and large digging claws. These fossils were dubbed "microsegnosaurids" and were present in the literature for years before the more detailed examination of the fossils proved that these bizzare therizinosaurs were too bizarre to be real.
Microsegnosauridae, a group of small, insectivorous maniraptors was one of the stranger radiations of Therizinosauria. Evolving in the Oligocene, migrosegnosaurids became quite common during the Miocene and Pliocene, when they spread across Eurasia.
Several well-preserved skeletons show small, lightly-built creatures with long, tubular snouts and powerful forelimbs. The retroverted pubis of the therizinosaurs was, with the microsegnosaurs, greatly reduced and pulled forward, like that of a basal coelurosaur, while the legs were reletively long and slender.
The obvious explanation for these adaptations is that these dinosaurs were ant-eaters, although exactly why ants were so tempting as a food source that the therizinosaurs could completely abandon their former lifestyle and favor of their consumption is unknown.
Formicornithoides europeanus[]
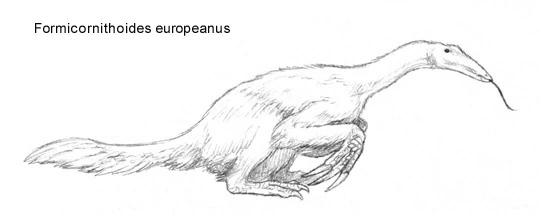

The early Oligocene microsegnosaurid Formicornithoides europeanus, which grew up to lengths of 3 to 5 meters long, is the first known species of myrmicophagous therizinosaurs. When the fragmentary skull and limb material the species is known from was first discovered, it couldn't be indisputably identified as a therizinosaurid. It wasn't before the discovery of Microsegnosaurus myrmicophagus that the animal could be properly classified.
Monstrengornis novaeaustralodromeus[]
A large, flightless enantiornithe from Australia in the Oligo/Miocene. It resembled HE's dromornids, but it evolved from an partridge like ancestor, being a large member of a linage of fowl like allospiziformes known as Australomentulaornidae, which ranged from partridge sized flying species to cassowary sized flightless ones, and then this. It was an herbivore, using its huge beak to break nuts and to break the legs of any dromeosaur that was near it.
EXTINCTION AT SPEC'S OLIGOCENE/MIOCENE TRANSITION (c. 24,000,000 - 23,000,000 b.p.)[]
The solar system is a messy place, littered with rocky and icy leftovers from its coalescence. The Earth suffers chronic encounters with these wanderers, and while most are too small to be of much consequence, a few, like the monster of RL's K-T, leave their scar upon both the face of the planet and the creatures that dwell upon it. At the very end of the Oligocene, such an event seems to have occurred.
Close to 23 million years ago, a great flaming rock fell from the sky. It slammed into the arctic and left a steaming crater 20 km across. It was not the largest object to have crossed Earth's path, nor was the impact the only one of its time. But for life on Earth, the rock had found the worst possible place to crash into.
The area surrounding the impact was undoubtedly devastated immediately. Thick clouds of ejecta in the air would have stalled photosynthesis across much of the Northern Hemisphere, breaking down food chains across North America and Eurasia and leading to the starvation of millions of animals. As devastating as this catastrophe must have been, the scorching of the sky was not the most serious consequence of the collision. The blast of the impact had, after vaporizing the surface sediments, encountered bedrock rich in anhydrite (calcium sulphate) and various carbonates. Great quantities of this material, some from over 1.7 km deep within the Earth, was thrown into the atmosphere where it lead to an increase in greenhouse gases and deadly falls of acid rain.
For the most part, the Southern Hemisphere seemed to escape any serious damage, although the last of South America's sauropods vanished at this time. Throughout North America and Eurasia, however, the dinosaurs were in peril. In North American fossil-bearing deposits produced at that time, one often finds as few as two or three dinosaur species where once there were over twenty.
On the whole, larger dinosaurs were more seriously affected than smaller ones. The last of the giant lambeosaurs along with most of the larger ceratopsians, eurolophs and neohadrosaurs vanished. Tyrannosaurs survived as a group, but lost many of their more bizarre members. The survivors were the errosaurs---small and generalised like the early cenoceratopsians and megahadrids. Without competition, these little predators quickly spread and proliferated in the Early Miocene.
On both Spec and, RL the eroded scar of the 23 million year-old impact, known as the Haughton crater, sits in the high arctic of North America (75°22'N, 89°41'W). The effects of the impact on RL's Cenozoic biosphere is still being investigated and it has been implicated in a number of regional extinctions. What is certain however, is that no megafaunal extinction event of the magnitude of Spec's decimated northern dinosaur faunas occurred in our native timeline. Why this would be the case when all physical aspects of the impact appear identical on both worlds is one of many issues under scientific scrutiny. One possibility concerns the lack of insulatory structures (such as fur or feathers) on most herbivorous dinosaurs. This may have made them more sensitive to such a sudden large-scale climate change compared to their mammalian RL counterparts.
SPEC'S MIOCENE EPOCH (c.24,000,000 - 5,000,000 b.p.)[]


The longest epoch of the Cenozoic bore witness to great changes in the geography and biota of the planet. As the wandering landmass of India finally collided with the Asian mainland, an event which spawned the highest mountains in the world, the Americas pushed westwards against the Pacific Plate, creating up the Rockies and the Andes. As Africa drew ever closer to Europe, it sounded the death knell for the ancient Tethys Sea. Cut off from neighbouring oceans, this once great ocean quickly evaporated into a vast dry salt-encrusted basin. While the Atlantic ultimately broke through the straights of Gibraltar and gave rise to the Mediterranean, the land link between Africa and Eurasia was to remain unbroken to the present day.
Vegetation on the land was lush and vibrant. Dense forests thrived across the globe. As more water was trapped at the poles,however, open grasslands and savannah spread rapidly, bringing about the evolution of some new clades while restricting and decimating others.
For life in the northern hemisphere, it was time of tremendous inter-continental migration and expansion. Africa, Eurasia, and North America were united, allowing free passage to wandering dinosaurs. At the start of the Miocene, the dominant Eurasian herbivores were the few euroloph families that had survived the Haughton impact. These natives were, however, soon locked in a losing ecological battle against successive waves of invading duckbills from neighbouring continents. Predatory dinosaurs also marched across the land bridges, the scaly African priscataurs rampaged across tropical Asia while Eurasian deinonychosaurs moved into Africa.
While Africa and the North experienced a mixing of faunas, the rest of the world was a study in isolation. Australia and South America remained worlds apart from any other landmass, surrounded by oceans on all sides. Both continents were enjoying a boom in ornithopod evolution that gave rise to a host weird endemic forms, from giant pseudosauropods to vicious rhynchoraptors. Australia began to lose much of its ancient rainforests as it charged northwards into drier latitudes.
Archaeopristrix stromeri []
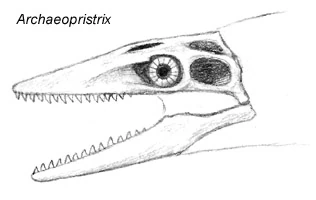
Growing up to lengths of four meters long, Archaeopristrix stromeri was the first known "fully modern" mosark and represents the best-preserved example of post-Cretaceous mosasaur diversity. Known from a virtually complete skeleton, Archaeopristrix is surprisingly similar to small mosarks like the sakhala and shows all the major developements that characterize today's lizardwhales, including a bent, fin-supporting tail, reduced hind flippers, and nostrils placed above the eyes.
Life in the seas was rich and varied. Giant predatory saurocetes and sharks prowled the tropics. while large swimming birds taking advantage of the rich polar waters. At several points during the Miocene, the the seas experienced sharp drops in temperature, accompanied by an expansion of the Antarctic ice sheet. However, these episodes were not severe enough to trigger anything more that mild, localised extinctions at sea or on land.
Tricerolophus chooi[]
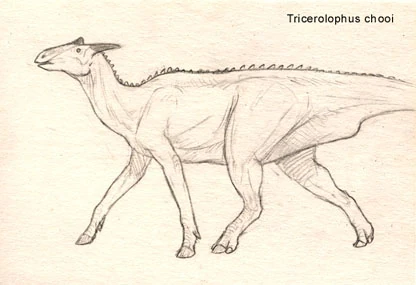
Growing up to lengths of seven meters long, Tricerolophus chooi is the first ungulaped hadrosaur known from Europe, dating back to middle Miocene. It may well be the common ancestor, or at least close relative, of many later Eurasian hadrosaurs. Unlike the modern saurolopes of Africa, Tricerolophus had only three horns, but seems to have been a good quadrapedal runner.
Microsegnosaurus mymicophagus[]
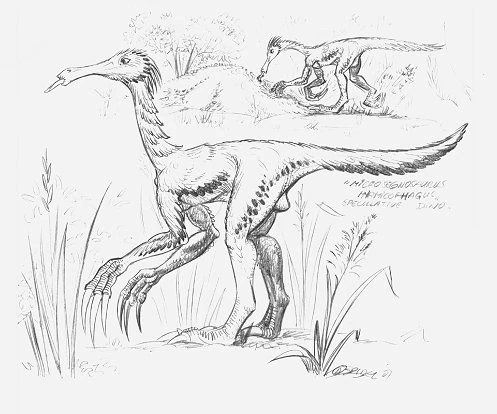
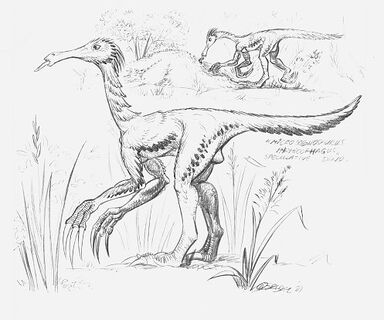
Reconstructions of Microsegnosaurus myrmicophagus, a pre-Ice Age therizinosaur which grew up to lengths of 2.5 meters long. Note the enlarged hands and elongated snout, both supposed to be adaptations to a myrmicivorous lifestyle.
Micronychus polyonyx[]
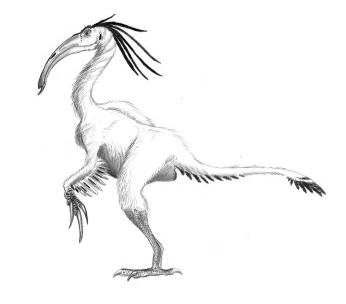
Scientists have debated the validity of the Microsegnosaurs due to some recent fossil discoveries in Eurasia which place the validity of this group of therizinosaurs in to question. This is mostly due to that some of these fossils turned out to be a jumbled mess of unknown of indeterminate therizinosaur remains and alvarezasaur remains. As of now, only a few species are considered to be valid, but even then this recent research could potentially make them dubious.
Recent studies of "microsegnosaurus", the first-discovered and most complete "microsegnosaurid" discovered proved that what was thought to be a single organism was, in fact, a chimera. The "ant-eating therizinosaur" may have actually been an amalgum of two unrelated organisms, a small therizinosaur possibly related to today's Seculasauridae and an alvarezsaur, the last known from the Eurasian continent. In the midst of this controversy, some work will be done to sort things out.
The three other species of "microsegnosaurid" (all of which based upon scattered elements) proved to be either therizinosaur of alvarezsaur material, lumped together with "microsegnosaurus" because of similarities with one part or another of its patch-work skeleton
Teratodon maturus[]
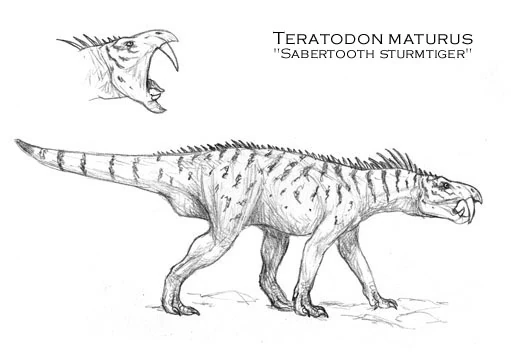
Growing up to lengths of four meters long, the Miocene Teratodon maturus is an early rhynchoraptoran megapredator. It isn't exacly clear why this animal became extinct, but competition from the more advanced rynchoraptorans, and the fact that Teratodon seems not to have been much of a runner may have played a part in it's demise. Several Teratodon skeletons have been found, including a mummified specimen show a row of sturmtiger-like spines on this animal's back run down to the base of the tail, which has gained Teratodon the nickname "sabertooth sturmtiger".
Wabulasaurus archeri[]
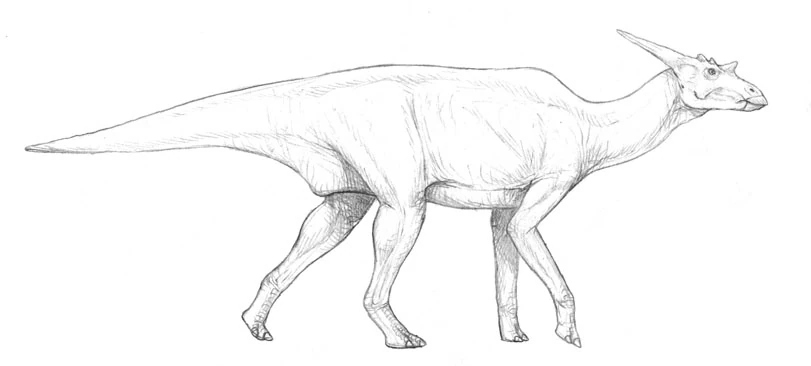

Ozpec's Riversleigh deposits (in northeastern Queensland) have produced the remains of a wide variety of fossil euclasaurs. The 5-million-year-old Wabulasaurus archeri is one the largest and most striking of these specimens. This 7 meter long, 2-ton animal appears to have been an early offshoot of the line leading to the living hexacorns. While generally similar to its modern-day relative, Wabulasaurus differed in the configuration of horns, its narrow beak, and the presence of large ceratopsian-like jugal processes. The postcranial skeleton is imperfectly known but appears to have been more gracile than the modern hexacorn.
Eomucrodontosaurus srnkai[]
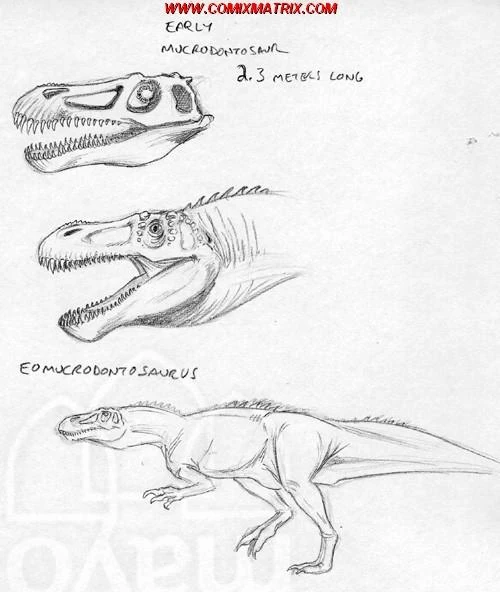
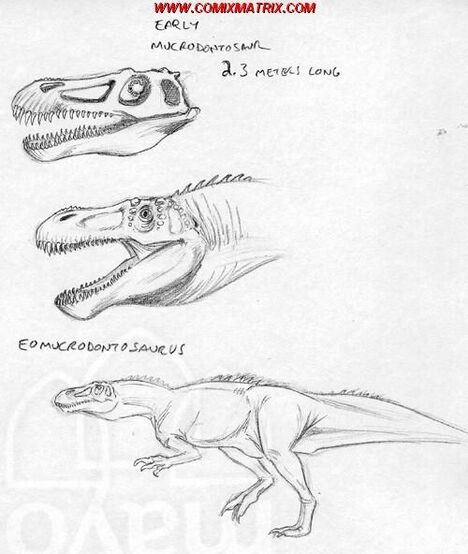
Here in this picture, you can see a reconstruction of Eomucrodontosaurus srnkai, a small carnivore from Madagascar which grew up to lengths of up to 2.3 meters, an early product of the abelisaurid radiation that produced the Mucrodontosaurus of today. Unlike the modern day Murcodontosaurus, this species doesn't appear to be as semi-aquatic as its modern day relative.
Mirificranium coronatus[]
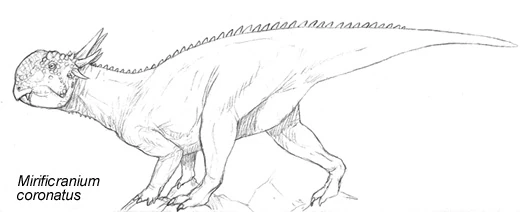
Both on HE and Spec the continent of Australia is known to host archaic, surprising and just plain weird forms of life. But to uncover some of the most perplexing creatures, you have to dig deep, as a group of specpaleontologists discovered. While hoping to uncover the ancestor of the rhynchoraptorans, they uncovered a startling specimen that didn't seem to make sense at all: a partial skull with fragmentary postrcania that looked like it could have belonged to a strange pachycephalosaur with a crown of spikes and two prominent fangs. The 2.6 meter long animal was named Mirificranium coronatus, meaning confusing crowned skull.
Such a find was hard swallow for several reasons. First of all, there were no known pachycephalosaur remains found in Australia prior to this, and secondly the age seemed too far removed from the last known Specworld pachycephalosaurs. Mirificranium could be dated to the Oligocene with reasonable accuracy, while the last of the pachycephalosaurs were supposed to have become extinct early Eocene.
An ulna is known from HE Australia that has been tentatively assigned to a neoceratopsian (possibly Leptoceratops), which has lead to some speculation that neoceratopsians may be of Gondwanan origin. Mirificranium coronatus was however the first ceratopsian to show up in the Australian fossil record, so if there is any connection between it and the Early Cretaceous ulna, there remains a question about where the Australian ceratopians were hiding all this time. The Mirificranium postcrania also hints that the body of the animal was probably not that of the typical neoceratopsian, but rather like that of the more basal ceratopians. If a recently discovered partial postcrania from Early Miocene turns out to belong to Stegocephalosaurus, then the stegocephalosaurs were most likely at least habitual bipeds.
Stegocephalosaurus baiamei[]

The tale got another twist when another, more complete skull was found from early Miocene deposits. This 2.9 meter long animal, Stegocephalosaurus baiamei (meaning creator god Baiame's roofheaded reptile) had a less flamboyant skull covered by a flat bony pad. Though there were enough similarities in the skull morphology to classify the animal as a marginocephalian, it didn't seem to belong to a pachycephalosaur after all. Instead it had a lot in common with the most basal ceratopsians!
There is still much more we don't know about stegocephalosaurs. We can only guess that they used their bony heads in intraspecific combat as pachycephalosaurs may have done in their time, but the purpose of the fangs is unclear. Another unanswered question is their fate. While they seem so rare in the fossil record that it is too early to make assumptions about when exactly they disappeared, they did not make it to the holocene. Their extinction is indeed a great shame, as from live stegocephalosaurs we might have learned a great deal about the evolution of marginocephalians both in our world as in spec.
Rhinolophosaurus aumalai[]
The massive rhinolophosaurs of the Oligocene and Miocene were the largest of their breed, strongly convergent with hadrosaurian ungulapeds. Indeed, rhinolophosaur material in France was first mistaken for very early ungulaped remains but the first complete rhinolophosaur skeleton, christened Rhinolophosaurus aumalai, was obviously not a hadrosaurian, and so the older finds, under scrutiny, turned out to be of the same stock. This massive creature grew up to lengths of 14 meters long and weight up to 6 tons.

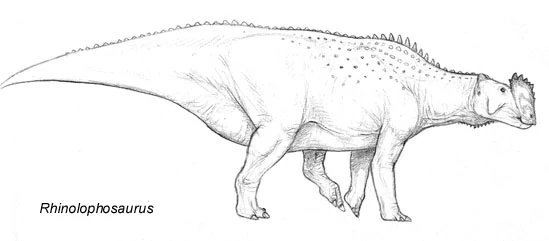
While the head of Rhinolophosaurus is indeed rather similar to a hadrosaur, with its battery of chewing teeth and bulbous nose, the rest of the body is not. The legs are short compared to the torso, and the neural spines over the shoulders are enlarged, traits found only in the truly huge hadrosaurs like hmungos and brutons. With such legs and spines, rhinolophs could only have been plodding, graviportal grazers, the earliest known in Eurasia. The tail, which in hadrosaurs counterbalances the body, is, as with all known rhinolophosaurs, devoid of stiffening tendons.
During their time, the rhinolophosaurs were the most successful herbivores in Eurasia, overshadowing the more familiar ceratopsians and hadrosaurs for the duration of the Oligocene epoch. The Haughton impact that began the Miocene took a heavy toll on the eurolophs, drastically reducing rhinolophosaur diversity and outright killing several other euroloph clades (about which very little is known). The rhinolophosaurs came back as strong as ever in the Miocene, producing such forms as the famous Rhinolophosaurus, still the most complete rhinolophosaur material found to date. Rhinolophosaurus and its kin continued quite happily up to the end of the Miocene, but the climatic changes of the Pliocene, coupled with the influx of African ungulapeds from the south spelt doom for the rhinolophosaurs as was the case with the larger Eurolophs.
Therizinonychus polynyx[]
- (Picture Needed)
Although all modern therizinosaurs are herbivores, in the not-too-distant past, their family tree included a predatory branch. Therizinonychus polynyx was the most common species of this extinct group. T. polynyx lived in Asia during the Miocene. Using its powerful body and large, sharp claws, Therizinonychus would overturn contemporary ankylosaurs, exposing their soft bellies. After ankylosaurs went into decline during the mid-Miocene, the therizinonychosaurs went with them. The clade's final species, such as the enormous Carnofalx bestialis, showed some tendancy toward a more varied diet, but died at the end of the Pliocene.
SPEC'S PLIOCENE EPOCH (c.5,000,000 - 1,700,000 b.p.)[]


The geography of the Pliocene Specworld looked quite similar to the present-day map. As North and South Ameirca finally met, the isthmus of Panama rose from the sea, separating the tropical Atlantic from the Pacific and ending South America's long isolation. The Mediterranean was fully formed, but was barred from the Indian Ocean by the Sinai peninsula, which joined Africa and Eurasia. A permanent polar icecap had started to form at the Arctic, shifting the warm Gulf Stream as cool currents flowed down the Labradore coast. Temperatures on land and in the sea continued to cool.
Many kinds of marine invertebrates vanished, unable to adapt to the changing temperatures, but were quickly replaced by more modern forms. The productive cool waters at the poles soon played host to a crowd of newly evolved animals taking advantage of their expanded habitat. Swimming birds radiated further and grew increasingly large. Enormous filter-feeding cephalopods harvested the clouds of krill. In warmer waters, new species of lizard-whales terrorised the oceans.
On land, the vegetation was shifting in response to these cooler, drier conditions. While trees still dominated the uplands, in the mid-continental belts, they were giving way to open grasslands. Low-lying areas once covered in green forests and woodlands were now transformed into endless seas of golden grass.
The rise of the grasslands lead to the loss of many ancient browsing herbivores. The armoured ankylosaurs, already uncommon since the Oligocene, declined to a handful of species in America and Australia. Most of the last of the eurolophs were gone, minus a few species who escaped the wrath of extinction, while only a single genus of high-browsing sauropod remained in Africa.
As some groups faded with the vanishing forests others rose to meet the evolutionary challenge. Throughout Africa and Asia, vast of herds of hoofed ungulapedes were on the march, these weird duckbills soon radiated into an incredible variety of forms. Cooler northern latitudes witnessed herbivorous niches becoming increasingly dominated by two of the most unlikely of candidates - the therizinosaurian theropods and the paraselenodont mammals. In North America, the neohadrosaur duckbills continued their monopoly of the large herbivore guilds. Also making their presence felt were the frill-necked ceratopsians. These dinosaurs had been absent from North America since the end of the Oligocene and their recent arrival from Asia heralded the return of giant horned herds to the continent.
Biogeographically, the most important event of the Pliocene was the linking of the two Americas. Much of the unique South American megafauna vanished in the face of North American immigrants. The northern dinosaurs had dealt with climatic change and faunal interchange with Asia throughout the Cenozoic, thus were full of competitive vigour. Their southern counterparts, on the other hand, have lived on a more stable, isolated world for millions of years and in most cases fare poorly against the invaders.
Soon, duckbills and ceratopsians had displaced many unique South American ornithopod families, while tyrannosaurs and hesperonychids had replaced the abelisaurs and endemic notoraptors, minus a few exceptions. Of the large southern herbivores, only the giant pseudosauropods held their own. However, the migrations were not one-way, as neotropical viriosaurs and marsupials flourished in the north.
The only major landmasses still completely isolated were Antarctica and Australia. The former was now virtually lifeless, but the latter boasted a staggering display of weird endemics. The thick forests that once covered most of the island continent were now restricted to the margins, giving way to open woodlands and savannah. One lineage of herbivores, the euclasaurs, had undergone an explosive evolutionary radiation akin to that of the Old World ungulapedes. Hunting these odd herbivores were their predatory rhynchoraptor cousins.
One of Spec's lesser-known dinosaur clades are the vanguards, America's ankylosaur-anologues, despite the fact that ankylosaurs still thrive, but their numbers have been reduced. Though these brutes have called NA their home since the Eocene, recent fossil finds have revealed that vanguards, or a closely related group lived in Asia from the Late Pliocene to the Mid-Pleistocene. Specpaleontologists are unsure of whether to place these fossils within Vanguardidae, so until a verdict can be reached, the fossils have been placed within their own group, Gojiramimidae. A number of features distinguish gojiramimids from the American vanguards, including powerful back legs, suggesting that they could've reared up on their hind legs for a few moments; a row of stegosaur-like spikes running down the end of their tails; and a series of (kaiju-esque) spines going down their backs.
Gojiramimids date back to Late Pliocene Russia with "Gojiramimus splendens", a 20-ft long low-browser. This species already possessed many of its descendents' notable features. Gojiramimid diversity remained at fairly decent levels, with fossils being found throughout Southeast Asia, until the Mid-Plesitocene, when the group started to face rough competition from the Formosicornids, which had just recently arrived from India and Tibet. It was from this point on that the Gojiramimids declined in number and diversity. One of the last members of the clad was "Gojiramimus sinensis" from Eastern China. It was thought that "G. sinensis" was the group's final member until the discovery of "G. japonicus", a sheep-sized descendant from Japan. It seems that "G. japonicus" thrived in it's island home until somewhere around 500,000 years ago, when it died out for unknown reasons.
Ayleeia breviops[]

Ayleeia breviops is an extinct North American ceratopsid, which grew up to the size same size, or possibly even larger, of the modern day Dinoceratops horridus. Though this animal lived in the Pliocene, it was surprisingly primitive, being very similiar to Late Cretaceous leptoceratopids. It lacked brow or nose horns, but seems to have had some kind of horny bumps on its neck frill. A recently discovered mummified specimen has revealed an interesting detail about its diet: much of its the stomach contents were made up of what seems to have been tree bark chewed up twigs and branches. How Ayleeia managed to digest such food is still unknown - or perhaps the individual in question was just starving and desperate enough to eat anything.
Projugalceratops problematicus []
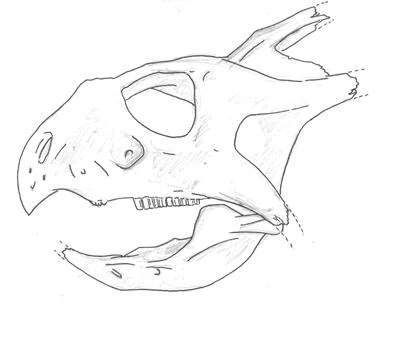
2.5 million year-old Projugalceratops problematicus, unearthed in Spec's Florida, is the only jugalceratopsine known from Spec's fossil record. This generalized herbivore was quite similar to Cretaceous Protoceratops and the present-day dawnhorn of Asia, but its affinities to other jugalceratopsines are uncertain. Most paleontologists had assumed the jugalceratopsines evolved from a dinoceratops -like ancestor, but the generalized Projugalceratops is more similar to Cretaceous Protoceratops and the present-day dawnhorn of Asia than to any dinoceratopsine. It estimated this creautre grew up to lengths of 4 to 5 meters long.
To further complicate matters, this North American fossil bears a very strong resemblance to the poorly-understood Frill-tusker of the Amazon. Perhaps the frill-tusker and Projugalceratopsform a group ancestral to all other jugalceratopsines, but more specimens must be studied to determine whether the species' similarities are phylogenic or merely convergent.
Mokelesaurus gargouille[]

Though in the Holocene mokelesaurs (excluding the dwarf forms on isolated islands) have been restricted to the continent of Africa, they were once found in large parts of Europe and western Asia. The first mokelesaurs migrated to Europe with their terrestial titanosaur cousins in the Miocene, but these first forms show less adaptations to aquatic lifestyle than modern mokeles.
In the Pliocene European waterways were dominated by the now-extinct species, Mokelesaurus gargouille, which, before the ice ages, ranged from the British isles to the Persian gulf . It was little different from the modern Nile mokele except for its larger size. The largest known M.gargouille skeleton, found in southern France, indicates the species could reach the length of 16 meters. It is yet unknown whether M. gargouille evolved in Europe or in Africa, but it seems to have given rise to the Mediterranean minimokeles (Minimokelesaurus insularis).
Megabeluasaurus magnacephalus[]
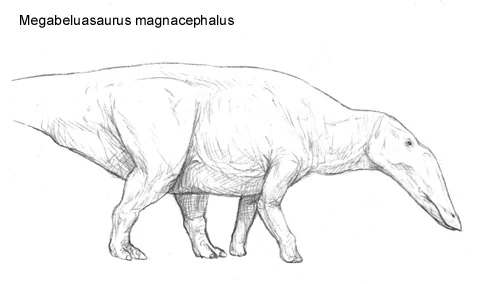
Megabeluasaurus magnacephalus was the largest and one of the last paramegahadrine species, even though it never surpassed the elephantine hmungos in size. Growing up to lengths of nine meters long. The first Megabeluasaurus remains found consisted of a nearly intact skull but very little postcranial material. Based on the enormous size of the skull, the animal was first reconstructed as an enormous hmungo-like grazer. Later finds revealed that the skull of Megabeluasaurus was actually very large in proportion to the rest of its body. The dinosaur seems to have been a dweller of the wetlands whose diet consisted mostly of aquatic plants very much like its cousin, the modern sludger.
Carnofalx bestialis []

Carnofalx bestialis was first thought to be one of the last and largest surviving members of therizinonychosaurs in the Pliocene. It turns out it was actually a 4 meter long oviraptorosaur. This heavily built animal was probably mainly a scavenger, but may have been partly omnivorous as well.
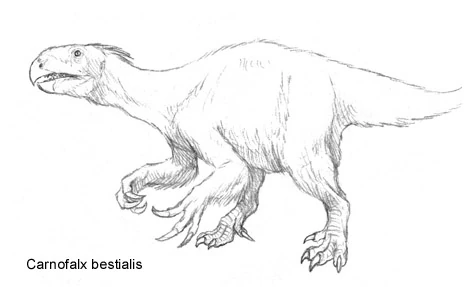
Ceratociraphus pervulgatus[]
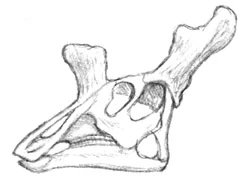
Ceratociraphus pervulgatus is an Pliocene ungulapede browser whose remains have been found in northern Africa, the middle east and India. Originally it was tentatively classified as an offshoot of Formosicornidae, but a more detailed analysis has revealed this animal to be a ciraphadrid. The unusual horn arrangements of Ceratociraphus may give an insight to how the horns of their formosicornid (and ultracornid) relatives may have evolved, although those of Ceratociraphus were blunt and massive, probably used in pusing or butting contests beteween males.
Hipposaurus (Hippohadrus) grandis[]
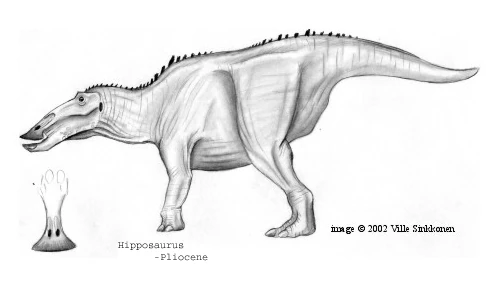
The last of the hippohadridae family was the mighty Hipposaurus, a ten-meter-long monster with powerful limbs and a bizzare skull. The head of Hipposaurus was deep, but very narrow until half-way down the snout, when the beak flared out into a spatula shape. Meanwhile, the lower jaw curves upward as it flares, forming a cutting edge with the upper mandible. The jaw is thus constructed in such a way that the dinosaur could only have been able to feed by holding its head perpendicular to the ground and dragging its beak along the turf like a giant spoon. Most paleontologists agree that these strange features were an adaptation for grazing upon early grasses.
As the Oligocene gave way to the Miocene, hipposaur diversity began to wane. By the Pliocene only the large and specialized Hipposaurus remained until it, too, succumbed to the changing climate and competition from other herbivores. A possible name change to "Hippohadrus grandis" has been proposed in order to not confuse it with the small therapsid which lived during the Permian period.
Pentacerolophus progenitor[]
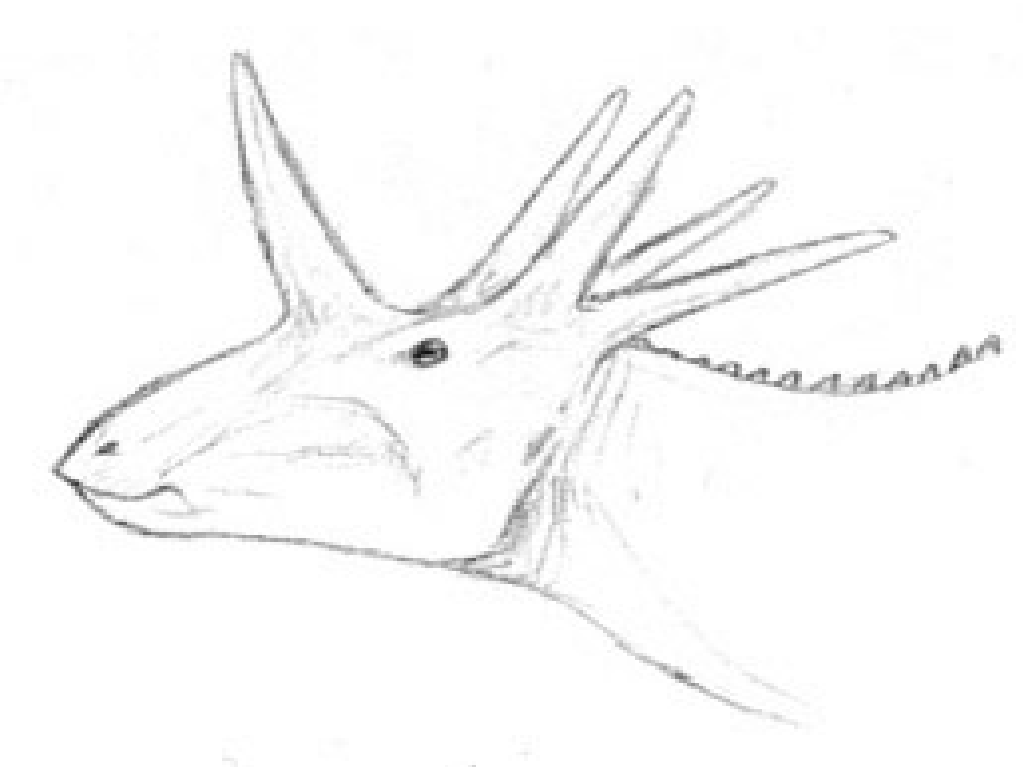

Pentacerolophus is an extinct genus that fits between ealines and catoblepids. This large grazer lived in Asia Minor where it evolved, and southern Europe from late Miocene to Middle Pliocene. Its appearance coincides with the beginning of the rhinolophosaurs' decline. Pentacerolophus primigenitus, the earliest known Pentacerolophus species from the late Miocene, was not much larger than modern ealines, at about 2 meters long, but the heavily built Pentacerolophus ingens was nearly twice the size of a yale which grew up to 4 meters long. Pentacerolophus disappears soon after the extinction of Rhinolophosaurus aumalai, apparently replaced by its progeny: the larger and more robust Catoblepas species and its close relatives. Pictured here is Pentacerolophus progenitor, the largest of all the species of Pentacerolophus which grew up to lengths of roughly 7 meters long.
Probactyrannus gracilis[]
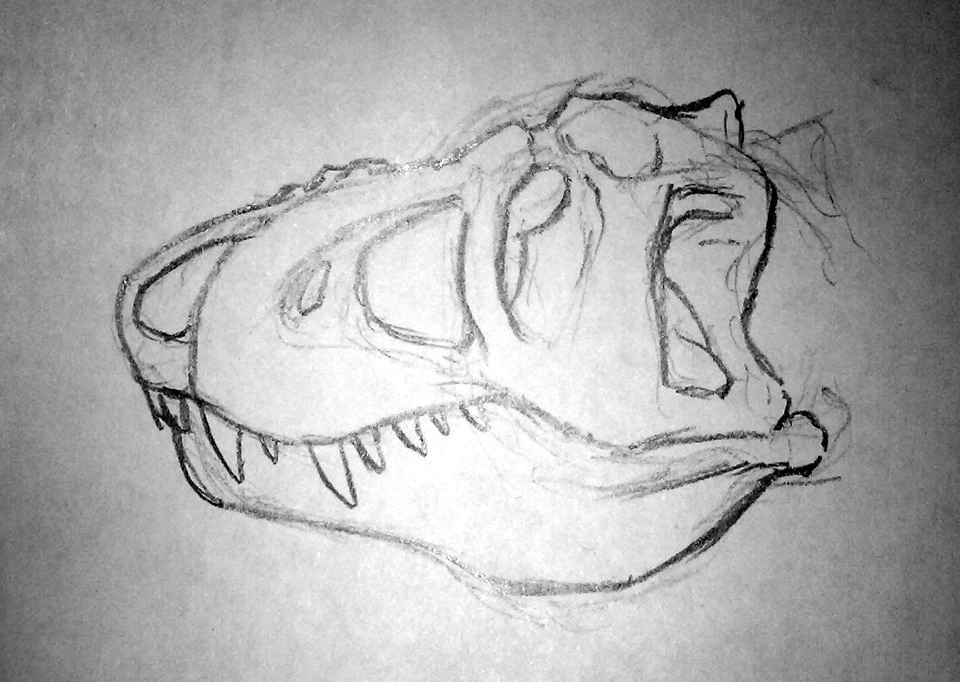
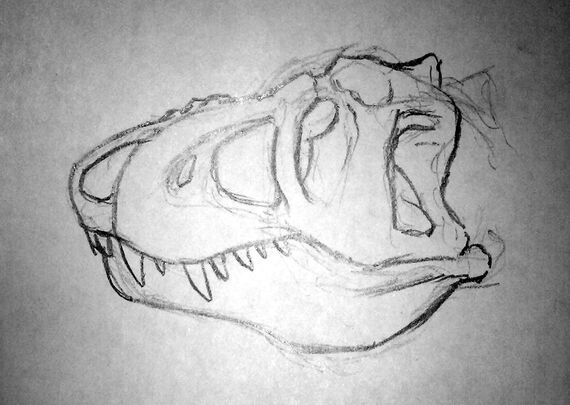
Probactyrannus gracilis (North America) Note: Seeing how the original artwork for Probactyrannus gracilis was lost, a new piece of artwork from its original artist, Christopher Srnka, has been recreated. All credit for the new artwork goes to Christopher Srnka.
Probactyrannus gracilis was a slender tyrannosauroid with elongated limbs and an enlarged nasal cavity. Dating from the middle Pliocene, this North American predator represents one of the many tyrannosauroid branches that met their untimely demise with the coming of the Ice Age.
SPEC'S QUATERNARY PERIOD (c.1,700,000 - 0 b.p.)[]


A little less than 2 million years ago, the Earth entered what can only be described as a climatic rollercoaster, a time which marked the end of the Neogene Period and the start of the Quaternary Period. Thus began a series of drastic cooling phases for the planet, the famous Ice Ages.
In RL, the Quaternary is divided into two epochs - the Pleistocene and the Holocene (the time from the last glaciation up until the present day). Although these units are sometimes used on Spec for purposes of convenience, it should be noted that:
- a) the biotic changes that have characterised the Holocene on RL are far less profound on Spec (see below)
- b) evidence collected on both worlds suggests that this current phase of mild climate is little more than a short interglacial period and that the Specworld is heading towards yet another round of glaciation. On RL the activities of industrialised humanity appears to be reversing this situation.
From about 1.7 million to 11,000 years ago, the world experienced at least four major glacial episodes. Great sheets of ice spread out from the Arctic, covering much of North America and Europe. Increased glaciation also occured in the southern hemisphere although to a much lesser extent. At its peak, about 44 million sq km of the planet's surface was ice-bound. With so much water locked away, sea-levels fell by over 100 metres, resulting in the devastation of local marine faunas and the creation of several land-bridges, including those across the Bering Strait, the English Channel, the Torres Strait, and the Sea of Japan. After a time, the ice would retreat and the world would be given the brief respite of an interglacial that saw the return of milder conditions.
The effect of the Ice Ages, both on the landscape and the creatures that inhabited it, was profound. The flow of the glaciers carved great valleys in the Earth, while accumulating boulder clays and moraines. Strongs winds near the ice sheets carried fine-grained dust to distant lands, resulting in thick loess deposits in Africa, central Asia, and midwestern North America.
With regards to the vegetation, the coming of the Ice Age heralded the arrival of vast areas tundra in the north, providing a rich feeding ground for cold-adapted grazers, such as polar therizinosaurs and large paraselenodont mammals. In the lower latitudes, some areas acquired lush woodlands while others faced increasing aridity. When the ice retreated, temperate forests returned to the high-latitudes, accompanied by herds of hadrosaurs. Each phase of glaciation and interglacial proved to be of benefit to some organsisms at the expense of others. The ranges of some animals expanded while other were reduced to scattered populations only to have the situation reversed when the extent of the ice shifted.
In all physical aspects, the progression of the Ice Ages of Spec appears to have been identical to that of RL. However, with regards to life on the planet, there is one startling contrast. The conclusion of the last Ice Age on RL broadly coincided with the extinction of many large terrestrial animals - mammoths, giant deer, ground sloths, sabre-tooth cats, etc. As far as can be determined, such a phenomenon did not occur on Spec. 11,000 years ago, the ranges of many tundra-dwelling dinosaurs, such as the arctotitan, were certainly greatly reduced, often accompanied by a reduction in body-size. Some areas also experienced localised extinctions, particularly North America, which lost its ceratopsians as great areas of woodland were converted to open praries. But on the whole, probably over 90% of all of Specworld's Ice Age megafaunal genera are still alive and well. This evidence points the finger of blame firmly at humanity for the extinction of the great beasts of our own home time-line. Spec, never subjected to the curse of the killer ape, remains the abode of giants.
Macropokemus robustus[]
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Macropokemus robustus, the "giant chilla" is known only from a few fragments of fossilized bone uncovered from Pleistocene deposits in Spec's China. These fossils suggest a very large creature, perhaps two meters in length, with a set of powerful, carnivorous-looking teeth. While modern chillas are quite harmless, this Ice Age mammal must have been a terror.
Smilotyrannus horridus[]

{kind=link}
Smilotyrannus horridus (Northern Eurasia and possibly Northern North America) (PLEASE IGNORE THE OLD TEXT)
Smilotyrannus horridus evolved to hunt the giant therizinosaurs during the last ice age, and grew up to 13 m long. It evolved huge "canine" teeth to bite through the thick plumage and fat of an extinct species of Arctotitan, its primary prey. Dinofuzz covered most of its body, including the toes. While this terrifying creature is now gone, its closest living relative, Smilotyrannus imperator, still thrives in the very same area that Smilotyrannus horridus thrived in.
Pseudocastor megadontis[]
A multituberculate which had entered the beaver niche, though it is the forward-side teeth (rather than the incisors) which are used. Fossils of these creautres date back to the Plestiocene epoch and is found in areas near the Black Swamp.
Paradoxadont megaincisor[]
Another species of extinct paraselodont with two large & long molars on either side of the upper jaw, with a long comb-like row of molars on each side of the lower jaw, with tusk-like sabers erupting from the front of the lower jaw. Also features a large & tall crest which runs from the eyes to the nose. possibly for fighting other males...or, like the antlers of deer and the horns of saurolope, as an extravagance to impress females. Fossils have been found in Eurasia and North America.
EXAMPLES OF THE EXTINCT FAUNA BASED ON THEIR RESPECTIVE TIME PERIOD[]
Cretaceous []
- Gigantala cranitus (Pterosaur)
- Gigantala hastarostris (Pterosaur)
- Baropecudis cornufrons (Pachycephalosaur)
- Spinocephalus superstes (Pachycephalosaur)
- Parasaurolophus palaeocaenicus (Hadrosaur)
- Antarctoptera hyperion (Pterosaur)
Paleocene[]
- Anserodromeus antiquus (Ungulaped)
- Helioraptor kaamos (Dromaeosaur)
- Archeotyrannus kaamos (Tyrannosaur)
Eocene[]
- Murgonocheirus australis (Pterosaur)
- Saltotyrannus nanus (Tyrannosaur)
- Protokagru aegyptiacum (Noasaur)
- Eomantarx termieri (Ankylosaur)
- Eohadrus excelsius (Hadrosaur)
- Megacaudia nameni (Hadrosaur)
- Brontoceratops robustus (Ceratopsian)
- Gigantops dominus (Ceratopsian)
- Paraxenoceratops oplos (Ceratopsian)
- Eobrachiceratops antiquus (Ceratopsian)
- Patriopokemus mordax (Pokemurid)
- Phobotyrannus robustus (Tyrannosaur)
- Orgetorix ozmandias (Tyrannosaur)
- Balaenanguis primitivus (Mosasaur)
- Arbroraptor mongoliensis (Dromaeosaur)
- Spinotitan grandis (Sauropod)
- Titanotarsus steveirwini (Sauropod)
- Pisciraptor nauticus (Spinosaur)
- Massosuchus europensis (Spinosaur)
- Carnosuchus tarbos (Spinosaur)
- Afrosauravis gracilis (Ornithomimid?)
- Rheamimus leptocauda (Ornithomimid?)
- Mehenotornis sternbergi (Ornithomimid?)
- Tarwii tarwii (Ornithomimid?)
- Struthiarctos robustus (Ornithomimid?)
- Struthiarctos africanus (Ornithomimid?)
- Errosaurus borealis (Tyrannosaur)
- Lycovenator messelensis (Deinonychosaur)
- Gruorostrum tingamarri (Deinonychosaur)
- Protocedunasaurus foxi (Cedunasaur)
- Protocedunasaurus messelensis (Cedunasaur)
Oligocene[]
- Inferanas cadaviperidis (Anseriforme)
- Protomoloch bonapartei (Abelisaur)
- Bavarionyx cursor (Eurolophia)
- Sauropodimimus giganticus (Hadrosaur)
- Gryphorhynchus robustus (Rhynchoraptor)
- Gryphorhynchus aenigmaticus (Rhynchoraptor)
- Paramegahadrus vetus (Hadrosaur)
- Procurvihadrus specensis (Hadrosaur)
- Protobrachioceratops gracilis (Cereatopsian)
- Arcotitian colossusis (Titanosaur)
- Aetiogenoi eletherios (Pterosaur)
- Formicornithoides europeanus (Microsegnosaur)
- Pygmihadrus mirus (Hipposaur)
- Bellasaurus leidyi (Hipposaur)
Miocene[]
- Monstrengornis novaeaustralodromeus (Allospiziforme)
- Archaeopristrix stromeri (Mosasaur)
- Tricerolophus chooi (Ungulaped)
- Teratodon maturus (Rhynchoraptor)
- Mirificranium coronatus (Stegocephalosaur)
- Stegocephalosaurus baiamei (Stegocephalosaur)
- Pikodon maximus (Torvodont)
- Micronychus polyonyx (Microsegnosaur)
- Microsegnosaurus myrmicophagus (Microsegnosaur)
- Eomucrodontosaurus srnkai (Noasaur)
- Taurosaurus gracilis (Ceratopsian)
- Megataurosaurus indica (Ceratopsian)
- Trooraptor negris (Rhynchoraptor)
- Therizinonychus polynyx (Therizinosauria)
- Pentacerolophus progenitor (Ungulaped)
- Wabulasaurus archeri (Euclasauria)
Pliocene[]
- Rhynchoraptor robustus (Rhynchoraptor)
- Ayleeia breviops (Dinoceratopsian)
- Projugalceratops problematicus (Dinoceratopsian)
- Carnofalx bestialis (Oviraptorosaur)
- Rhinolophosaurus aumalai (Rhinolophosaur)
- Megabeluasaurus magnacephalus (Hadrosaur)
- Ceratociraphus pervulgatus (Ungulaped)
- Pentacerolophus ingens (Ungulaped)
- Pentacerolophus primigenitus (Ungulaped)
- Neotitanosaurus uralensis aka the Russian Grassbag (Titanosaur)
- Mokelesaurus gargouille aka the European Mokele (Titanosaur)
- Hipposaurus grandis (Hipposaur)
- Probactyrannus gracilis (Tyrannosaur)
- Rhinocerocristatus giganteus (Pterosaur)
- Gojiramimus splendens (Vanguard)
- Nanodon marshalensis (Abelisaur)
Plestiocene[]
- Macropokemus robustus (Pokemurid)
- Monoceratops acutus (Cenoceratopsian)
- Neotitanosaurus ultramaximus aka Ultramax Grassbag (Titanosaur)
- Neotitanosaurus dispar (Titanosaur)
- Smilotyrannus horridus aka the Horrific Sabre-Tyrant (Tyrannosaur)
- Smilotyrannus silvensis (Tyrannosaur)
- Tetraketos moorei (Plesiosaur)
- Gojiramimus sinensis (Vanguard)
- Gojiramimus japonicus (Vanguard)
- Cornucantus belli yukonensis aka the Yukon Elumbe (Ungulipede)
- Epippiosaurus progenitor aka the American Torg (Hadrosaur)
- Neosimoedosaurus barbaricina (Champsosaur)
- Pseudocastor megadontis (Multituberculate)
- Paradoxadont megaincisor (Paraselodont)